Успіхи у вивченні нуклеїнових кислоті біосинтезу білка призвели до створення низки методів, що мають велике прикладне значення в медицині, сільському господарстві та інших галузях.
Після того, як був вивчений генетичний код та основні принципи зберігання та реалізації спадкової інформації, розвиток молекулярної біології зайшов у глухий кут, тому що не було методів, які дозволяли маніпулювати генами, виділяти та змінювати їх. Поява цих методів сталася у 1970-1980-х роках. Це дало потужний поштовх до розвитку цієї галузі науки, яка і сьогодні переживає період розквіту. Насамперед, ці методи стосуються отримання індивідуальних генів та їх введення у клітини інших організмів (молекулярне клонування та трансгенез, ПЛР), а також методів визначення послідовності нуклеотидів у генах (секвенування ДНК та РНК). Нижче ці методи будуть розглянуті докладніше. Ми почнемо з найпростішого базового методу - електрофорезу і потім перейдемо до складніших методів.
ЕЛЕКТРОФОРЕЗ ДНК
Це базовий спосіб роботи з ДНК, що застосовується разом з практичними всіма іншими способами для виділення необхідних молекул та аналізу результатів. Для поділу фрагментів ДНК за довжиною застосовується метод електрофорезу гелі. ДНК - кислота, її молекули містять залишки фосфорної кислоти, які відщеплюють протон і набувають негативного заряду (рис. 1).
Тому в електричному полімолекули ДНК рухаються до анода – позитивно зарядженого електрода. Це відбувається в розчині електролітів, що містить іони-носія заряду, завдяки чому цей розчин проводить струм. Щоб розділити фрагменти, застосовується щільний гель із полімерів (агарози або поліакриламіду). Молекули ДНК "заплутуються" в ньому тим більше, чим вони довші, і тому найдовші молекули рухаються найповільніше, а найкоротші - найшвидше (рис. 2). Завчасно або після електрофорезу гель обробляють барвниками, що зв'язуються з ДНК і флуоресціюють в ультрафіолетовому світлі, і отримують картину смуг у гелі (див. рис. 3). Для визначення довжин фрагментів ДНК зразка порівнюють їх з маркером - набором фрагментів стандартних довжин, нанесених паралельно на той же гель (рис. 4).



Найважливішими інструментами для роботи з ДНК є ферменти, що здійснюють перетворення ДНК у живих клітинах: ДНК-полімерази, ДНК-лігази та рестрикційні ендонуклеази, або рестриктази. ДНК-полімеразиздійснюють матричний синтез ДНК, що дозволяє розмножувати ДНК у пробірці. ДНК-лігазизшивають між собою молекули ДНК або заліковують проломи них. Рестрикційні ендонуклеази, або рестриктази, розрізають молекули ДНК за певними послідовностями, що дозволяє вирізати окремі фрагменти із загальної маси ДНК. Ці фрагменти можуть у деяких випадках містити окремі гени.
рестриктази
Послідовності, відомі рестриктазами, симетричні, і розриви можуть відбуватися в середині такої послідовності або зі зрушенням (в тому самому місці в обох нитках ДНК). Схема дії різних типів рестриктазу показана на рис. 1. У першому випадку виходять так звані «тупі» кінці, тоді як у другому – «липкі» кінці. У разі «липких» кінців дна ланцюг виявляється коротшим за інший, утворюється однониткова ділянка з симетричною послідовністю, однаковою на обох кінцях, що утворюються.
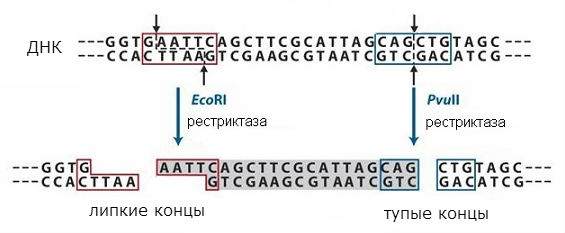
Кінцеві послідовності будуть однаковими при розщепленні будь-якої ДНК даною рестриктазою і можуть знову з'єднуватися, оскільки мають комплементарні послідовності. Їх можна пошити за допомогою ДНК-лігази та отримати єдину молекулу. Таким чином вдається об'єднати фрагменти двох різних ДНК та отримати так звані рекомбінантні ДНК. Цей підхід використовується у методі молекулярного клонування, що дозволяє отримати індивідуальні гени та ввести їх у клітини, які можуть утворювати закодований у гені білок.
молекулярне клонування
У молекулярному клонуванні використовується дві молекули ДНК - вставка, що містить ген, що цікавить, і вектор- ДНК, що у ролі носія. Вставку "вшивають" у вектор за допомогою ферментів, отримуючи нову, рекомбінантну молекулу ДНК, потім цю молекулу впроваджують у клітини-господарі, і ці клітини утворюють колонії на живильному середовищі. Колонія - це потомство однієї клітини, тобто клон, всі клітини колонії генетично ідентичні і містять одну й ту саму рекомбінантну ДНК. Звідси термін "молекулярне клонування", тобто отримання клону клітин, що містять фрагмент ДНК, що цікавить нас. Після того, як колонії, що містять вставку, що цікавить нас, отримані, можна різними методами характеризувати цю вставку, наприклад, визначити її точну послідовність. Також клітини можуть виробляти білок, що кодується вставкою, якщо вона містить функціональний ген.
При впровадженні рекомбінантної молекули клітини відбувається генетична трансформація цих клітин. Трансформація- процес поглинання клітиною організму вільної молекули ДНК із середовища проживання і вбудовування їх у геном, що зумовлює появі в такої клітини нових для неї успадкованих ознак, притаманних організму-донора ДНК. Наприклад, якщо вбудована молекула містить ген стійкості до антибіотика ампіциліну, то трансформовані бактерії зростатимуть у його присутності. До трансформації ампіцилін викликав їхню загибель, тобто у трансформованих клітин виникає нова ознака.
ВЕКТОРИ
Вектор повинен мати ряд властивостей:
По-перше, це відносно невелика молекула ДНК, щоб їй легко маніпулювати.
По-друге, для того, щоб ДНК зберігалася і розмножувалася в клітці, вона повинна містити певну послідовність, що забезпечує її реплікацію (початок реплікації, або origin of replication).
По-третє, вона має утримувати ген-маркерякий забезпечує відбір тільки тих клітин, в які потрапив вектор. Зазвичай це гени стійкості до антибіотиків - тоді в присутності антибіотика всі клітини, що не містять вектора, гинуть.
Клонування генів найчастіше проводять у клітинах бактерій, тому що вони прості у культивуванні та швидко розмножуються. У клітині бактерії зазвичай присутня одна велика кільцева молекула ДНК, завдовжки кілька мільйонів пар нуклеотидів, що містить усі необхідні бактерії гени - бактеріальна хромосома. Крім неї в деяких бактеріях існують невеликі (кілька тисяч пар нуклеотидів) кільцеві ДНК, які називаються плазмідами(Рис. 2). Вони, як і основна ДНК, містять послідовність нуклеотидів, що забезпечує здатність ДНК реплікуватися (ori). Плазміди реплікуються незалежно від основної (хромосомної) ДНК, тому присутні у клітині у великій кількості копій. Багато з таких плазмід несуть гени стійкості до антибіотиків, що дозволяє відрізнити клітини, що несуть плазміду, від звичайних клітин. Найчастіше використовуються плазміди, що несуть два гени, що забезпечують стійкість до двох антибіотиків, наприклад, до тетрацикліну та апміциліну. Існують прості методи виділення таких плазмідних ДНК, вільних від ДНК основної хромосоми бактерії.

ЗНАЧЕННЯ ТРАНСГЕНЕЗУ
Перенесення генів з одного організму в інший називають трансгенезом, а такі модифіковані організми - трансгенними. Методом перенесення генів у клітини мікроорганізмів отримують рекомбінантні білкові препарати для потреб медицини, зокрема, людські білки, що не викликають імунного відторгнення - інтерферони, інсулін та інші білкові гормони, клітинні фактори росту, а також білки для виробництва вакцин. У більш складних випадках, коли модифікація білків проходить правильно тільки в клітинах еукаріотів, застосовують трансгенні клітинні культури або трансгенних тварин, зокрема, худобу (насамперед кіз), яку виділяє необхідні білки в молоко, або ж білки виділяють з їхньої крові. Так отримують антитіла, фактори зсідання крові та інші білки. Методом трансгенезу отримують культурні рослини, стійкі до гербіцидів та шкідників та які мають інші корисні властивості. За допомогою трансгенних мікроорганізмів очищають стічні води та борються із забрудненнями, існують навіть трансгенні мікроби, які можуть розщеплювати нафту. Крім цього, трансгенні технології незамінні в наукових дослідженнях- розвиток біології сьогодні немислимо без рутинного застосування методів модифікації та перенесення генів.
технологія молекулярного клонування
вставки
Для отримання індивідуального гена з якогось організму з нього виділяють усю хромосомну ДНК і розщеплюють її однією або двома рестриктазами. Ферменти підбирають так, щоб вони не розрізали цікавий для нас ген, а робили розриви по його краях, а в плазмідної ДНК робили 1 розрив в одному з генів стійкості, наприклад, до ампіциліну.
Процес молекулярного клонування включає такі етапи:
Розрізання та зшивання - конструювання із вставки та вектора єдиної рекомбінантної молекули.
Трансформація – впровадження рекомбінантної молекули у клітини.
Селекція – відбір клітин, які отримали вектор із вставкою.
розрізання та зшивання
Плазмідну ДНК обробляють тими самими рестриктазами, і вона перетворюється на лінійну молекулу, якщо підібрано таку рестриктазу, яка вносить у плазміду 1 розрив. У результаті кінцях всіх утворюються фрагментів ДНК виявляються одні й самі липкі кінці. При зниженні температури ці кінці з'єднуються випадковим чином і їх зшивають ДНК-лігазою (див. рис. 3).

Отримують суміш кільцевих ДНК різного складу: деякі з них будуть містити певну послідовність ДНК хромосомної ДНК, сполучену з бактеріальною ДНК, інші - разом з'єднані фрагменти хромосомної ДНК, а треті - відновлену кільцеву плазміду або її димер (Рис. 4).

трансформація
Далі цією сумішшю проводять генетичну трансформаціюбактерій, які не містять плазміди. Трансформація- процес поглинання клітиною організму вільної молекули ДНК із середовища проживання і вбудовування їх у геном, що зумовлює появі в такої клітини нових для неї успадкованих ознак, притаманних організму-донора ДНК. У кожну клітину може проникнути та розмножитися там лише одна плазміда. Такі клітини поміщають на тверде живильне середовище, в якому міститься антибіотик тетрациклін. Клітини, які не потрапила плазміда, цьому середовищі не зростатимуть, а клітини, що несуть плазміду, утворюють колонії, у кожному з яких перебувають нащадки лише однієї клітини, тобто. всі клітини в колонії несуть ту саму плазміду (див. рис. 5).

Селекція
Далі стоїть завдання виділити тільки клітини, в які потрапив вектор із вставкою, і відрізнити їх від клітин, що несуть лише вектор без вставки або зовсім не несуть вектора. Цей процес відбору потрібних клітин називається селекцією. Для цього застосовують селективні маркери- зазвичай гени стійкості до антибіотиків у складі вектора, та селективні середовища, що містять антибіотики або інші речовини, що забезпечують селекцію
У намі прикладі клітини з колоній, що виросли в присутності ампіциліну, пересівають на два середовища: у першій є ампіцилін, а в другій – тетрациклін. Колонії, що містять тільки плазміду, зростуть на обох середовищах, а колонії, у плазмідах яких знаходиться вбудована хромосомна ДНК на середовищі з тетрацикліном, не зростуть (рис. 5). Серед них спеціальними методами відбирають ті, які містять цікавий для нас ген, вирощують у достатніх кількостях і виділяють плазмідну ДНК. З неї за допомогою тих же рестриктаз, які використовувалися при отриманні рекомбінантної ДНК, вирізують індивідуальний ген, що цікавить. ДНК цього гена може використовуватися для визначення послідовності нуклеотидів, введення в якийсь організм для отримання нових властивостей або синтезу потрібного білка. Такий метод виділення генів називається молекулярним клонуванням.
ФЛУОРЕСЦЕНТНІ БІЛКИ
Як гени-маркери при дослідженнях еукаріотичних організмів дуже зручно використовувати флуоресцентні білки. Ген першого флуоресцентного білка, зеленого білка, що флуорескує (green fluorescent protein, GFP)був виділений з медузи Aqeuorea victoria і впроваджений у різні модельні організми (див. рис. 6). У 2008 році О. Сімомура, М. Чалфі та Р. Тсьєн отримали Нобелівську премію за відкриття та застосування цього білка.

Потім було виділено гени інших флуоресцентних білків - червоного, синього, жовтого. Ці гени були модифіковані штучно, щоб отримати білки з корисними властивостями. Різноманітність флуоресцентних білків показано на рис. 7 де зображена чашка Петрі з бактеріями, що містять гени різних флуоресцентних білків.

застосування флуоресцентних білків
Ген флуоресцентного білка можна зшивати з геном будь-якого іншого білка, тоді при трансляції утворюватиметься єдиний білок - трансляційно злитий білок, або фьюжн(fusion protein), який флуоресціює. Таким чином можна вивчати, наприклад, локалізацію (розташування) будь-яких білків, що цікавлять, в клітині, їх переміщення. За допомогою експресії флуоресцентних білків тільки у певних типах клітин можна помічати клітини цих типів у багатоклітинному організмі (див. рис. 8 – мозок миші, в якому окремі нейрони мають різні кольори за рахунок певної комбінації генів флуоресцентних білків). Флуоресцентні білки – незамінний інструмент сучасної молекулярної біології.

ПЛР
Ще один метод отримання генів називається полімеразною ланцюговою реакцією (ПЛР). В його основі лежить здатність ДНК-полімераз добудовувати другу нитку ДНК за комплементарною ниткою, як це відбувається в клітинах при реплікації ДНК.
Точки початку реплікації у цьому методі задаються двома невеликими фрагментами ДНК, які називаються затравками,або праймерами. Ці затравки комплементарні кінцям гена, що цікавить, на двох ланцюгах ДНК. Спочатку хромосомну ДНК, з якої треба виділити ген, змішують із затравками і нагрівають до 99 про С. Це призводить до розриву водневих зв'язків та розходження ниток ДНК. Після цього температуру знижують до 50-70 про З (залежно від довжини та послідовності затравок). У цих умовах затравки приєднуються до комплементарних ділянок хромосомної ДНК, утворюючи правильну подвійну спіраль (див. рис. 9). Після цього додають суміш усіх чотирьох нуклеотидів, необхідних для синтезу ДНК, та ДНК-полімеразу. Фермент подовжує затравки, будуючи двоспіральну ДНК місця прикріплення затравок, тобто. від кінців гена, до кінця одноланцюгової хромосомної молекули. 
Якщо тепер знову нагріти суміш, то хромосомні та знову синтезовані ланцюги розійдуться. Після охолодження до них знову приєднаються затравки, які беруться у великій надлишку (див. рис. 10).

На знову синтезованих ланцюгах вони приєднаються не до того кінця, з якого починався перший синтез, а до протилежного, так як ланцюги ДНК антипаралельні. Тому у другому циклі синтезу на таких ланцюгах добудується лише послідовність, що відповідає гену (див. рис. 11).

В даному методі застосовується ДНК-полімераза з термофільних бактерій, здатна витримувати кип'ятіння і працююча при температурах 70-80 про С, її не треба додавати щоразу, а достатньо внести на початку досвіду. Повторюючи процедури нагрівання та охолодження в тій же послідовності, ми можемо в кожному циклі подвоювати число послідовностей, обмежених з двох кінців затравки (див. рис. 12).

Після приблизно 25 таких циклів кількість копій гена збільшиться більш ніж мільйон разів. Такі кількості легко можна відокремити від внесеної в пробірку хромосомної ДНК і використовувати для різних цілей.
секвенування ДНК
Ще одним важливим досягненням є розробка методів визначення послідовності нуклеотидів у ДНК. секвенування ДНК(Від англ. Sequence – послідовність). Для цього необхідно отримати чисті з інших ДНК гени одним з описаних методів. Потім ланцюга ДНК розділяють нагріванням і додають до них затравку, мічену радіоактивним фосфором або флуоресцентною міткою. Зверніть увагу, що береться одне затравлення, комплементарне одного ланцюга. Потім додається ДНК полімераза та суміш з 4-х нуклеотидів. Така суміш ділиться на 4 частини і до кожної додається один з нуклеотидів, модифікований так, що третій атом дезоксирибози він не містить гідроксильної групи. Якщо такий нуклеотид включиться в синтезований ланцюг ДНК, його подовження зможе продовжуватися, т.к. полімеразі нікуди приєднуватиме наступний нуклеотид. Тому синтез ДНК після включення такого нуклеотиду обривається. Таких нуклеотидів, званих дидезоксинуклеотиди, додається значно менше, ніж звичайних, тому урвище ланцюга відбувається лише зрідка і в кожному ланцюгу в різних місцях. В результаті виходить суміш ланцюгів різної довжини, на кінці кожної з них стоїть той самий нуклеотид. Таким чином довжина ланцюга відповідає номеру нуклеотиду в послідовності, що вивчається, наприклад, якщо у нас був аденіловий дидезоксинуклеотид, а отримані ланцюги мали довжину 2, 7 і 12 нуклеотидів, значить в гені в другій, сьомій і дванадцятій позиції стояв аденін. Отриману суміш ланцюгів легко розділити за розмірами за допомогою електрофорезу, а синтезовані ланцюги виявити за радіоактивністю на рентгенівській плівці (див. рис. 10).

Виходить картина, наведена внизу малюнка, що називається радіоавтографом. Рухаючись по ньому знизу вгору і читаючи букву над колонками кожної зони ми отримаємо послідовність нуклеотидів, наведену малюнку праворуч від автографа. Виявилося, що синтез зупиняється не тільки дидезоксинуклеотидами, а й нуклеотидами, у яких у третьому положенні цукру приєднується якась хімічна група, наприклад флюоресцентний барвник. Якщо кожен нуклеотид позначити своїм барвником, то зони, які отримують при розділенні синтезованих ланцюгів, світитимуться різним світлом. Це дозволяє проводити реакцію в одній пробірці одночасно для всіх нуклеотидів і розділяючи отримані ланцюги за довжиною, ідентифікувати кольори за нуклеотидами (див. рис. 11).

Такі методи дозволили визначити послідовності як окремих генів, а й прочитати цілі геноми. В даний час розроблені ще швидші методи визначення послідовностей нуклеотидів у генах. Якщо паровий геном людини був розшифрований великим міжнародним консорціумом з використанням першого наведеного методу за 12 років, другий, з використанням другого, за три роки, то це може бути зроблено за місяць. Це дозволяє передбачати схильність людини до багатьох захворювань та заздалегідь вживати заходів, щоб уникнути їх.
Розвиток біохімії, біофізики, генетики, цитохімії, багатьох розділів мікробіології та вірусології приблизно на початок 40-х років XX ст. впритул підвело до вивчення життєвих явищ на молекулярному рівні. Успіхи, досягнуті цими науками, одночасно і з різних сторін призвели до усвідомлення того факту, що саме на молекулярному рівні функціонують основні керуючі системи організму і що подальший прогрес цих наук залежатиме від розкриття біологічних функцій молекул, що становлять тіла організмів, їх участі у синтезі та розпаду, взаємних перетвореннях та репродукції сполук у клітині, а також обміну енергією та інформацією, що відбувається при цьому. Так на стику цих біологічних дисциплін з хімією та фізикою виникла зовсім нова галузь – молекулярна біологія.
На відміну від біохімії, увага сучасної молекулярної біології зосереджена переважно на вивченні структури та функції найважливіших класів біополімерів – білків та нуклеїнових кислот, перші з яких визначають саму можливість протікання обмінних реакцій, а другі – біосинтез специфічних білків. Зрозуміло тому, що чітке розмежування молекулярної біології та біохімії, відповідних розділів генетики, мікробіології та вірусології неможливо.
Виникнення молекулярної біології було тісно пов'язане з розробкою нових методів дослідження, про які вже йшлося у відповідних розділах. Поряд із розвитком електронної мікроскопії та інших методів мікроскопічної техніки велику роль відіграли розроблені у 50-х роках методи фракціонування клітинних елементів. Вони ґрунтувалися на вдосконалених методах диференціального центрифугування (А. Клод, 1954). До цього часу вже були досить надійні способи виділення та фракціонування біополімерів. Сюди належить, зокрема, запропонований А. Тизеліусом (1937; Нобелівська премія, 1948) метод фракціонування білків за допомогою електрофорезу, методи виділення та очищення нуклеїнових кислот (Е. Кей, А. Даунс, М. Севаг, А. Мирський та ін. ). Паралельно у багатьох лабораторіях світу розроблялися різні методи хроматографічного аналізу (А. Мартін та Р. Сінг, 1941; Нобелівська премія, 1952), згодом суттєво вдосконалені.
Неоціненну послугу у розшифровці структури біополімерів зіграв рентгеноструктурний аналіз. Основні принципи рентгеноструктурного аналізу було розроблено у Королівському коледжі Лондонського університету під керівництвом У. Брегга групою дослідників, куди входили Дж. Бернал, А. Лондсдейл, У. Астбері, Дж. Робертсон та інших.
Слід особливо відзначити дослідження професора Московського державного університету А. Р. Кізеля з біохімії протоплазми (1925 – 1929), що мали найважливіше значення для подальшого становлення молекулярної біології. Кизель завдав удару міцно укоренився уявленню, що в основі будь-якої протоплазми лежить особливе білкове тіло - пластин, що нібито визначає всі її найважливіші структурні та функціональні особливості. Він показав, що пластини - це білок, який зустрічається тільки у міксоміцетів, і то на певній стадії розвитку, і що жодного постійного компонента - єдиного скелетного білка - у протоплазмі не існує. Тим самим вивчення проблеми будови протоплазми та функціональної ролі білків вийшло на правильний шлях і одержало простір для свого розвитку. Дослідження Кізеля завоювали світове визнання, стимулювавши вивчення хімії складових частин клітини.
Термін "молекулярна біологія", вперше використаний англійським кристалографом професором Лідського університету У. Астбері, виник, ймовірно, на початку 40-х років (до 1945 р.). Основні рентгеноструктурні дослідження білків і ДНК, проведені Астбері в 30-х роках, послужили основою для подальшого успішного розшифрування вторинної структури цих біополімерів. У 1963 р. Дж. Бернал писав: "Пам'ятник йому буде встановлено всією молекулярною біологією - наукою, яку він назвав і справді заснував" * , У літературі цей термін з'явився вперше, мабуть, у 1946 р. у статті У. Астбері "Прогрес рентгеноструктурного" аналізу органічних та фібрилярних сполук", опублікованій в англійському журналі "Природа"**. У своїй Гарвіївській лекції Астбері (1950) відзначав: "Мені приємно, що зараз термін молекулярна біологія вже досить широко вживається, хоча мало ймовірно, що я першим запропонував його. Він мені подобався і я вже давно намагався його поширювати". Вже в 1950 р. Астбері було ясно, що молекулярна біологія має справу насамперед із структурою та конформацією макромолекул, вивчення яких має вирішальне значення для розуміння функціонування живих організмів.
* (Biogr. Mem. Fellows Roy. Soc, 1963, v. 9, 29.)
** (W. T. Astbury. Progress of X-ray analysis of organic and fibre structures.- Nature,. 1946, v. 157, 121.)
*** (W. T. Astbury. Adventures in Molecular Biology. Thomas Springfield, 1952, p. 3.)
Перед молекулярною біологією стояли і стоять, власне, ті самі завдання, що і перед усією біологією в цілому, - пізнання сутності життя та його основних явищ, зокрема таких, як спадковість та мінливість. Сучасна молекулярна біологія насамперед покликана розшифрувати структуру та функцію генів, шляхи та механізми реалізації генетичної інформації організмів на різних стадіях онтогенезу та на різних етапах її зчитування. Вона покликана розкрити тонкі механізми регуляції активності генів та клітинної диференціювання, з'ясувати природу мутагенезу та молекулярні основи еволюційного процесу.
Встановлення генетичної ролі нуклеїнових кислот
Для становлення молекулярної біології найбільше значення мали такі відкриття. У 1944 р. американські дослідники О. Евері, К. Мак-Леод (Нобелевська премія, 1923) і М. Мак-Карті показали, що виділені з пневмококів молекули ДНК мають трансформуючу активність. Після гідролізу цих ДНК дезоксирибонуклеазою їхня трансформуюча активність повністю зникала. Тим самим було вперше було переконливо доведено, що генетичними функціями у клітині наділена саме ДНК, а чи не білок.
Заради справедливості слід зазначити, що явище бактеріальної трансформації було виявлено значно раніше відкриття Евері, Мак-Леода та Мак-Карті. У 1928 р. Ф. Гріффіт опублікував статтю, в якій повідомив, що після додавання до невірулентних (некапсульованих) пневмококів убитих клітин капсульованого вірулентного штаму одержувана суміш клітин стає згубною для мишей. Більш того, живі клітини пневмококів, що виділяються із заражених цією сумішшю тварин, були вже вірулентними і мали полісахаридну капсулу. Тим самим у цьому досвіді було показано, що під впливом якихось компонентів убитих клітин пневмококів некапсульована форма бактерій перетворюється на капсулоутворюючу вірулентну форму. Через 16 років Евері, Мак-Леод і Мак-Карті замінили в цьому досвіді вбиті цілі клітини пневмококів їх дезоксирибонуклеїновою кислотою і показали, що саме ДНК має трансформуючу активність (див. розділи 7 і 25). Значення цього відкриття важко переоцінити. Воно стимулювало вивчення нуклеїнових кислот у багатьох лабораторіях світу та змусило сконцентрувати увагу вчених саме на ДНК.
Поруч із відкриттям Эвери, Мак-Леода і Мак-Карти на початку 50-х вже накопичилося досить багато прямих і непрямих даних у тому, що нуклеїнові кислоти грають виняткову роль життєдіяльності і несуть генетичну функцію. На це, зокрема, вказував і характер локалізації ДНК у клітині та дані Р. Вендрелі (1948) про те, що вміст ДНК на клітину строго постійно і корелює зі ступенем плідності: у гаплоїдних статевих клітинах ДНК вдвічі менше, ніж у диплоїдних соматичних. На користь генетичної ролі ДНК свідчила також її виражена метаболічна стабільність. На початку 50-х років накопичилося багато різноманітних фактів, що свідчили, що більшість відомих мутагенних факторів діють переважно на нуклеїнові кислоти і, особливо, на ДНК (Р. Хочкісс, 1949; Г. Ефруссі-Тейлор, 1951; Е. Фріз , 1957 та ін.).
Особливого значення у встановленні генетичної ролі нуклеїнових кислот мало вивчення різних фагів та вірусів. У 1933 р. Д. Шлезінгер знайшов ДНК у бактеріофазі кишкової палички. З моменту виділення У. Стенлі (1935, Нобелівська премія, 1946) вірусу тютюнової мозаїки (ВТМ) у кристалічному стані розпочався новий етап у вивченні рослинних вірусів. У 1937 – 1938 рр. Співробітники Ротамстедської сільськогосподарської станції (Англія) Ф. Боуден і Н. Пірі показали, що багато виділених ними рослинних вірусів є не глобулінами, а являють собою рибонуклеопротеїди і містять як обов'язковий компонент нуклеїнову кислоту. На самому початку 40-х років були опубліковані роботи Г. Шрамма (1940), П. А. Агатова (1941), Г. Міллера та У. Стенлі (1941), що свідчили про те, що помітна хімічна модифікація білкового компонента не призводить до втрати інфекційності ВТМ. Це вказувало на те, що білковий компонент не може бути носієм спадкових властивостей вірусу, як продовжували вважати багато мікробіологів. Переконливі докази на користь генетичної ролі нуклеїнової кислоти (РНК) у рослинних вірусів були отримані у 1956 р. Г. Шраммом у Тюбінгені (ФРН) та X. Френкель-Конратом у Каліфорнії (США). Ці дослідники практично одночасно і незалежно один від одного виділили з ВТМ РНК і показали, що саме вона, а не білок, має інфекційність: у результаті зараження рослин тютюну цієї РНК у них відбувалося формування та розмноження нормальних вірусних частинок. Це означало, що РНК містить інформацію для синтезу та збирання всіх вірусних компонентів, у тому числі і вірусного білка. У 1968 р. І. Р. Атабеков встановив, що білок грає істотну роль при зараженні рослин - природою білка визначається спектр рослин-господарів.
У 1957 р. Френкель-Конрат вперше здійснив реконструкцію ВТМ із складових його компонентів – РНК та білка. Поряд із нормальними частинками він отримав змішані "гібриди", у яких РНК була від одного штаму, а білок - від іншого. Спадковість таких гібридів повністю визначалася РНК, і потомство вірусів належало до того штаму, РНК якого було використано отримання вихідних змішаних частинок. Пізніше досліди А. Гірера, Г. Шустера та Г. Шрамма (1958) та Г. Вітмана (1960 – 1966) показали, що хімічна модифікація нуклеїнового компонента ВТМ призводить до появи різноманітних мутантів цього вірусу.
У 1970 р. Д. Балтімор і Р. Темін встановили, що перенесення генетичної інформації може відбуватися як від ДНК до РНК, а й навпаки. Вони виявили у деяких онкогенних РНК-вірусів (онкорнавіруси) особливий фермент, так звану зворотну транскриптазу, який здатний на ланцюгах РНК комплементарно синтезувати ДНК. Це велике відкриття дозволило зрозуміти механізм вбудовування в геном господаря генетичної інформації РНК-вірусів і по-новому поглянути на природу їх онкогенної дії.
Відкриття нуклеїнових кислот та вивчення їх властивостей
Термін нуклеїнових кислот був введений німецьким біохіміком Р. Альтманом в 1889 р., після того, як ці сполуки були відкриті в 1869 р. швейцарським лікарем Ф. Мішером. Мішер екстрагував клітини гною розведеною соляною кислотою протягом кількох тижнів та отримав у залишку майже чистий ядерний матеріал. Цей матеріал він вважав характерною речовиною клітинних ядер і назвав його нуклеїном. За своїми властивостями нуклеїн різко відрізнявся від білків: він був більш кислим, не містив сірку, зате в ньому було багато фосфору, він добре розчинявся в лугах, але не розчинявся в розведених кислот.
Результати своїх спостережень над нуклеїном Мішер направив Гоппе-Зейлеру для опублікування в журналі. Описана ним речовина була настільки незвичайною (тоді з усіх біологічних сполук фосфору був відомий тільки лецитин), що Гоппе-Зейлер не повірив досвідам Мішера, повернув йому рукопис і доручив своїм співробітникам М. Плошу та Н. Любавіну перевірити його висновки на іншому матеріалі. . Робота Мішера "Про хімічний склад клітин гною" побачила світ двома роками пізніше (1871). У той же час були опубліковані роботи Гоппе-Зейлера та його співробітників щодо складу клітин гною, еритроцитів птахів, змій та інших клітин. Протягом наступних трьох років нуклеїн був виділений із тварин клітин та дріжджів.
У своїй роботі Мішер зазначав, що детальне вивчення різних нуклеїнів може призвести до встановлення відмінностей між ними, передбачивши цим ідею специфічності нуклеїнових кислот. Досліджуючи молоко лосося, Мішер встановив, що нуклеїн знаходиться в них у вигляді солі і пов'язаний з основним білком, який він назвав протаміном.
У 1879 р. в лабораторії Гоппе-Зейлер вивченням нуклеїнів почав займатися А. Коссель. У 1881 р. він виділив з нуклеїну гіпоксантин, однак у той час він ще сумнівався у походженні цієї основи та вважав, що гіпоксантин може бути продуктом деградації білків. У 1891 р. серед продуктів гідролізу нуклеїну Коссель виявив аденін, гуанін, фосфорну кислоту та ще одну речовину з властивостями цукру. За дослідження з хімії нуклеїнових кислот Косселю у 1910 р. було присуджено Нобелівську премію.
Подальші успіхи у розшифровці структури нуклеїнових кислот пов'язані з дослідженнями П. Левіна та співробітників (1911 – 1934). У 1911 р. П. Левін та В. Жакобс ідентифікували вуглеводний компонент аденозину та гуанозину; вони встановили, що до складу цих нуклеозидів входить D-рибоза. У 1930 р. Левін показав, що вуглеводним компонентом дезоксирибонуклеозидів є 2-дезокси-D-рибоза. З його робіт стало відомо, що нуклеїнові кислоти побудовані з нуклеотидів, тобто фосфорильованих нуклеозидів. Левін вважав, що основним типом зв'язку в нуклеїнових кислотах (РНК) є 2", 5"-фосфодіефірний зв'язок. Це уявлення виявилося помилковим. Завдяки роботам англійського хіміка А. Тодда (Нобелевська премія, 1957) та його співробітників, а також англійських біохіміків Р. Маркхема та Дж. Сміта на початку 50-х років стало відомо, що основним типом зв'язку в РНК є 3", 5"- фосфодіефірний зв'язок.
Левін показав, що різні нуклеїнові кислоти можуть відрізнятися за природою вуглеводного компонента: одні містять цукор дезоксирибозу, інші - рибозу. Крім того, зазначені два типи нуклеїнових кислот розрізнялися за природою однієї з основ: у нуклеїнових кислотах пентозного типу містився урацил, а в нуклеїнових кислотах дезоксипентозного типу – тимін. Дезоксипентозну нуклеїнову кислоту (за сучасною термінологією, дезоксирибонуклеїнова кислота – ДНК) зазвичай легко виділяли у великих кількостях із тимусу (зобної залози) телят. Тому вона отримала назву тимонуклеїнової кислоти. Джерелом нуклеїнової кислоти пентозного типу (РНК) служили головним чином дріжджі та зародки пшениці. Цей тип часто називали дріжджовою нуклеїновою кислотою.
На початку 30-х років досить міцно укоренилося уявлення, ніби для рослинних клітин характерна нуклеїнова кислота дріжджового типу, а тимонуклеїнова кислота властива лише ядрам тваринних клітин. Два типи нуклеїнових кислот - РНК і ДНК - тоді називали відповідно рослинної і тваринної нуклеїновими кислотами. Проте, як показали ранні дослідження А. Н. Білозерського, такий поділ нуклеїнових кислот є невиправданим. У 1934 р. Білозерський вперше виявив тимонуклеїнову кислоту в рослинних клітинах: з проростків гороху він виділив та ідентифікував тімін-піримідинову основу, характерну саме для ДНК. Потім він виявив тімін і в інших рослинах (насінні сої, квасолі). У 1936 р. А. Н. Білозерський та І. І. Дубровська виділили препаративно ДНК із проростків кінського каштана. Крім того, серія робіт, виконаних у 40-х роках в Англії Д. Девідсоном із співробітниками, переконливо показала, що рослинна нуклеїнова кислота (РНК) міститься у багатьох тваринних клітинах.
Широке застосування розробленої Р. Фельгеном і Г. Розенбеком (1924) цитохімічної реакції на ДНК та реакції Ж. Браше (1944) на РНК дозволило досить швидко та однозначно вирішити питання про переважну локалізацію цих нуклеїнових кислот у клітині. Виявилося, що ДНК зосереджена в ядрі, тоді як РНК концентрується переважно в цитоплазмі. Пізніше було з'ясовано, що РНК міститься як у цитоплазмі, так і в ядрі, а крім того, були виявлені цитоплазматичні ДНК.
Що стосується питання про первинну структуру нуклеїнових кислот, то до середини 40-х років у науці міцно утвердилося уявлення П. Левіна, згідно з яким усі нуклеїнові кислоти побудовані за одним типом і складаються з однакових так званих тетрануклеотидних блоків. У кожному з цих блоків, на думку Левіна, міститься чотири різні нуклеотиди. Тетрануклеотидна теорія будови нуклеїнових кислот значною мірою позбавляла ці біополімери специфічності. Тому не дивно, що всю специфіку живого пов'язували тоді лише з білками, природа мономерів яких набагато різноманітніша (20 амінокислот).
Першу пролом теоренуклеотидного будови нуклеїнових кислот пробили аналітичні дані англійського хіміка Дж. Гуланда (1945 - 1947). При визначенні складу нуклеїнових кислот азоту підстав він не отримав еквімолярного співвідношення підстав, як це мало б бути згідно з теорією Левіна. Остаточно тетрануклеотидна теорія будови нуклеїнових кислот впала в результаті досліджень Е. Чаргаффа та його співробітників (1949 – 1951). Для поділу основ, що вищеплюються з ДНК внаслідок її кислотного гідролізу, Чаргафф використовував хроматографію на папері. Кожна з цих підстав була точно визначена спектрофотометрично. Чаргафф помітив значні відхилення від еквімолярного співвідношення підстав у ДНК різного походження і вперше виразно заявив, що ДНК має виражену видову специфічність. Тим самим було покінчено з гегемонією концепції про специфічність білка у живій клітині. Аналізуючи ДНК різного походження, Чаргафф відкрив і сформулював унікальні закономірності складу ДНК, що увійшли до науки під назвою правил Чаргаффа. Згідно з цими правилами, у всіх ДНК, незалежно від походження, кількість аденіну дорівнює кількості тиміну (А = Т), кількість гуаніну дорівнює кількості цитозину (Г = Ц), кількість пуринів дорівнює кількості піримідинів (Г + А = Ц + Т), кількість основ з 6-аміногрупами дорівнює кількості основ з 6-кетогруп-пами (А+Ц=Г+Т). Разом про те, попри такі суворі кількісні відповідності, ДНК різних видів відрізняються за величиною відношення А+Т:Г+Ц. В одних ДНК кількість гуаніну та цитозину переважає над кількістю аденіну та тиміну (ці ДНК Чаргафф назвав ДНК ГЦ-типу); інші ДНК містили аденіну та тиміну більше, ніж гуаніну та цитозину (ці ДНК були названі ДНК АТ-типу). Отримані Чаргаффом дані щодо складу ДНК зіграли виняткову роль молекулярної біології. Саме вони лягли в основу відкриття будови ДНК, зробленої в 1953 Дж. Уотсоном і Ф. Криком.
Ще в 1938 р. У. Астбері і Ф. Белл за допомогою рентгеноструктурного аналізу показали, що площини основ у ДНК повинні бути перпендикулярними до довгої осі молекули і нагадувати як би стос пластин, що лежать один над одним. У міру вдосконалення техніки рентгеноструктурного аналізу до 1952 – 1953 рр. накопичилися відомості, що дозволили судити про довжину окремих зв'язків та кутах нахилу. Це дозволило з найбільшою ймовірністю уявити характер орієнтації кілець пентозних залишків у сахарофосфатном кістяку молекули ДНК. У 1952 р. С. Фарберг запропонував дві умоглядні моделі ДНК, які представляли складену або закручену саму на себе однотяжку молекулу. Не менш спекулятивна модель будови ДНК була запропонована у 1953 р. Л. Полінгом (лауреат Нобелівської премії, 1954) та Р. Корі. У цій моделі три закручені ланцюги ДНК утворювали довгу спіраль, стрижень якої було представлено фосфатними групами, а підстави розташовувалися зовні від нього. До 1953 М. Вілкінс і Р. Франклін отримали більш чіткі рентгеноструктурні картини ДНК. Їхній аналіз показав повну неспроможність моделей Фарберга, Полінга та Корі. Використовуючи дані Чаргаффа, зіставляючи різні поєднання молекулярних моделей окремих мономерів та дані рентгеноструктурного аналізу, Дж. Вотсон і Ф. Крик у 1953 р. дійшли висновку, що молекула ДНК має бути двотяжкою спіраллю. Правила Чаргаффа різко обмежили кількість можливих упорядкованих поєднань основ у запропонованій моделі ДНК; вони підказали Уотсону і Крику, що у молекулі ДНК має бути специфічне спарювання основ - аденіну з тиміном, а гуаніну з цитозином. Іншими словами аденіну в одному ланцюзі ДНК завжди суворо відповідає тиміну в іншому ланцюзі, а гуаніну в одному ланцюзі обов'язково відповідає цитозин в інший. Тим самим Уотсон і Крик вперше сформулювали виняткову важливість принцип комплементарної будови ДНК, згідно з яким один ланцюг ДНК доповнює іншу, тобто послідовність підстав одного ланцюга однозначно визначає послідовність підстав в іншому (комплементарному) ланцюгу. Стало очевидно, що у самій структурі ДНК закладено потенційну можливість її точного відтворення. Ця модель будови ДНК нині є загальновизнаною. За розшифровку структури ДНК Крику, Вотсону та Вілкінсу в 1962 р. було присуджено Нобелівську премію.
Слід зазначити, що ідея про механізм точного відтворення макромолекул та передачу спадкової інформації зародилася в нашій країні. У 1927 р. Н. К. Кольцов висловив припущення, що при розмноженні клітин відбувається репродукція молекул шляхом точного автокаталітичного відтворення наявних материнських молекул. Щоправда, тоді Кольцов наділяв цим властивістю не молекули ДНК, а молекули білкової природи, про функціональному значенні яких тоді нічого було відомо. Проте сама думка про автокаталітичне відтворення макромолекул та механізм передачі спадкових властивостей виявилася пророчою: вона стала керівною ідеєю сучасної молекулярної біології.
Проведені в лабораторії А. Н. Білозерського А. С. Спіріним, Г. Н. Зайцевою, Б. Ф. Ванюшиним, С. О. Урисон, А. С. Антоновим та іншими багаторічними дослідженнями (1957-1974) складу ДНК у самих різноманітних організмів повністю підтвердили закономірності, виявлені Чаргаффом, та повну відповідність з молекулярною моделлю будови ДНК, запропонованою Вотсоном та Криком. Ці дослідження показали, що ДНК різних бактерій, грибів, водоростей, актиноміцетів, вищих рослин, безхребетних і хребетних мають специфічність складу. Особливо різко відмінності у складі (змісті АТ-пар основ) виражені у мікроорганізмів, виявляючись важливою таксономічною ознакою. У вищих рослин та тварин видові варіації у складі ДНК виражені значно слабкіше. Але це зовсім не означає, що ДНК у них менш специфічна. Крім складу підстав специфічність переважно визначається їх послідовністю в ланцюгах ДНК.
Поряд із звичайними основами у складі ДНК та РНК були виявлені додаткові азотисті основи. Так, у складі ДНК рослин та тварин Г. Уайт (1950) знайшов 5-метилцитозин, а Д. Данн та Дж. Сміт (1958) виявили в деяких ДНК метильований аденін. Довгий час метилцитозин вважався характерною рисою генетичного матеріалу вищих організмів. У 1968 р. А. Н. Білозерський, Б. Ф. Ванюшин та Н. А. Кокуріна встановили, що він може зустрічатися також і в ДНК бактерій.
У 1964 р. М. Голд та Дж. Хурвітц відкрили новий клас ферментів, що здійснюють природну модифікацію ДНК – її метилювання. Після цього відкриття стало ясно, що мінорні (що містяться в малих кількостях) основи виникають вже на готовому полінуклеотидному ланцюзі ДНК у результаті специфічного метилювання залишків цитозину та аденіну в особливих послідовностях. Зокрема, за даними Б. Ф. Ванюшина, Я. І. Бур'янова та А. Н. Білозерського (1969) метилювання аденіну в ДНК кишкової палички може відбуватися в термінуючих кодонах. За даними А. Н. Білозерського та співробітників (1968 – 1970), а також М. Мезельсона (США) та В. Арбера (Швейцарія) (1965 – 1969) метилювання надає молекулам ДНК унікальні індивідуальні риси та у поєднанні з дією специфічних нуклеаз є частиною складного механізму, який здійснює контроль за синтезом ДНК у клітині. Іншими словами, характер метилювання тієї чи іншої ДНК визначає питання про те, чи вона може розмножуватися в даній клітині.
Практично в той же час почалося виділення та інтенсивне вивчення ДНК-метилаз та рестрикуючих ендонуклеаз; у 1969 – 1975 рр. встановлені нуклеотидні послідовності, відомі в ДНК деякими з цих ферментів (X. Бойєр, X. Сміт, С. Лін, К. Муррей). При гідролізі різних ДНК рестрицирующим ферментом вивіваються досить великі фрагменти з однаковими "липкими" кінцями. Це дає можливість як аналізувати структуру генів, як це зроблено в невеликих вірусів (Д. Натанс, З. Адлер, 1973 - 1975), а й конструювати різні геноми. З відкриттям цих специфічних ферментів рестрикції генетична інженерія стала відчутною реальністю. Вбудовані у невеликі плазмідні ДНК гени різного походження вже легко вводять у різні клітини. Так, отримано новий тип біологічно активних плазмід, що дають стійкість до деяких антибіотиків (С. Коен, 1973), введені рибосомальні гени жаби та дрозофіли в плазміди кишкової палички (Дж. Морроу, 1974; X. Бойєр, Д. Хогнесс, Р. , 1974 – 1975). Таким чином, відкриті реальні шляхи для отримання принципово нових організмів шляхом введення та вбудовування у їх генофонд різноманітних генів. Це відкриття може бути спрямоване на благо людства.
У 1952 р. Г. Уайт та С. Коен виявили, що в ДНК Т-парних фагів міститься незвичайна основа – 5-оксиметилцитозин. Пізніше з робіт Е. Волькіна та Р. Сінсхеймера (1954) та Коена (1956) стало відомо, що залишки оксиметилцитозину можуть бути повністю або частково глюкозидовані, внаслідок чого молекула фагової ДНК виявляється захищеною від гідролітичної дії нуклеаз.
На початку 50-х років з робіт Д. Данна та Дж. Сміта (Англія), С. Заменхофа (США) та А. Вакера (ФРН) стало відомо, що в ДНК можуть включатися багато штучних аналогів основ, заміщаючи іноді до 50% тиміну. Як правило, ці заміщення призводять до помилок при реплікації, транскрипції ДНК та трансляції та до появи мутантів. Так, Дж. Мармур (1962) встановив, що у ДНК деяких фагів замість тиміну міститься оксиметилурацил. У 1963 р. І. Такахаші та Дж. Мармур виявили, що в ДНК одного з фагів замість тиміну міститься урацил. Таким чином, звалився ще один принцип, за яким раніше поділяли нуклеїнові кислоти. З часів робіт П. Левіна вважалося, що характерною ознакою ДНК є тимін, а РНК – урацил. Стало ясно, що ця ознака не завжди надійна, і важливою відмінністю хімічної природи двох типів нуклеїнових кислот, як це представляється на сьогоднішній день, є лише характер вуглеводного компонента.
Під час вивчення фагів було розкрито багато незвичайних ознак організації нуклеїнових кислот. З 1953 р. вважалося, що це ДНК є двотяжкі лінійні молекули, а РНК - лише однотяжні. Це становище суттєво похитнулося в 1961 р., коли Р. Сінсхеймер виявив, що ДНК фага φ X 174 представлена однотяжкою кільцевою молекулою. Щоправда, потім з'ясувалося, що у такій формі ця ДНК існує лише у вегетативної фагової частинки, а реплікативна форма ДНК цього фага також є двотяжкою. Крім того, дуже несподіваним виявилося, що РНК деяких вірусів може бути двотяжкою. Цей новий тип макромолекулярної організації РНК було виявлено у 1962 р. П. Гоматосом, І. Таммом та іншими дослідниками у деяких вірусів тварин та у вірусу ранової пухлини рослин. Нещодавно В. І. Агол та А. А. Богданов (1970) встановили, що крім лінійних молекул РНК існують також замкнені або циклічні молекули. Циклічна двотяжка РНК виявлена ними, зокрема, у вірусу енцефаломієлокардиту. Завдяки роботам X. Дево, Л. Тіноко, Т. І. Тихоненко, Е. І. Будовського та інших (1960 – 1974) стали відомі основні риси організації (укладання) генетичного матеріалу у бактеріофагів.
Наприкінці 50-х років американський вчений П. Доті встановив, що при нагріванні відбувається денатурація ДНК, що супроводжується розривом водневих зв'язків між парами основ та розбіжністю комплементарних ланцюгів. Цей процес носить характер фазового переходу на кшталт "спіраль-клубок" і нагадує плавлення кристалів. Тому процес теплової денатурації ДНК Доті назвав плавленням ДНК. При повільному охолодженні відбувається ренатурацпя молекул, тобто возз'єднання комплементарних половинок.
Принцип ренатурації в 1960 р. був використаний Дж. Мармуром і К. Шильдкраутом для визначення ступеня "гібридизування" ДНК різних мікроорганізмів. Згодом Е. Болтон та Б. Мак-Карті удосконалили цей прийом, запропонувавши метод так званих ДНК-агарових колонок. Цей метод виявився незамінним у вивченні ступеня гомології нуклеотидної послідовності різних ДНК та з'ясуванні генетичної спорідненості різних організмів. Відкрита Доти денатурація ДНК у поєднанні з описаною Дж. Манделем та А. Херши* (1960) хроматографією на метильованому альбуміні та центрифугуванням у градієнті щільності (метод розроблений у 1957 р. М. Мезельсоном, Ф. Сталем та Д. Виноградом) розділення, виділення та аналізу окремих комплементарних ланцюгів ДНК Так, наприклад, В. Шибальськи (США), використовуючи ці прийоми для поділу ДНК лямбда фага, показав у 1967 - 1969 рр., що генетично активними є обидва ланцюжки фага, а не одна, як це було прийнято рахувати (С. Спігельман, 1961). Слід зазначити, що вперше ідея про генетичну значущість обох ланцюжків ДНК лямбда фага була висловлена в СРСР С. Є. Бреслером (1961).
* (За роботи з генетики бактерій та вірусів А. Херші спільно з М. Дельбрюком та С. Луріа були удостоєні 1969 р. Нобелівської премії.)
Для розуміння організації та функціональної активності геному першорядне значення має визначення нуклеотидної послідовності ДНК. Пошуки методів такого визначення проводять у багатьох лабораторіях світу. У М. Бір зі співробітниками з кінця 50-х років намагається встановити послідовність ДНК за допомогою електронної мікроскопії, але поки що безуспішно. На початку 50-х років з перших робіт Сінсхеймера, Чаргаффа та інших дослідників з ферментативної деградації ДНК стало відомо, що різні нуклеотиди в молекулі ДНК розподілені хоч і нехаотично, але нерівномірно. За даними англійського хіміка К. Бартона (1961), піримідини (їх понад 70%) зосереджені переважно у вигляді відповідних блоків. А. Л. Мазін і Б. Ф. Ванюшин (1968 - 1969) встановили, що різні ДНК мають різний ступінь зблоченості піримідинів і що в ДНК тварин організмів вона помітно зростає в міру переходу від нижчих до вищих. Отже, еволюція організмів відбито у структурі їх геномів. Саме тому для розуміння еволюційного процесу в цілому порівняльне вивчення структури нуклеїнових кислот набуває особливого значення. Аналіз структури біологічно важливих полімерів і насамперед ДНК вкрай важливий і для вирішення багатьох приватних питань філогенетики та таксономії.
Цікаво відзначити, що англійський фізіолог Е. Ланкестер, який вивчав гемоглобіни молюсків, що рівно 100 років тому передбачив ідеї молекулярної біології, писав: "Хімічні відмінності різних видів та пологів тварин і рослин мають таке ж важливе значеннядля з'ясування історії їх походження, як і розбіжності у тому формі. Якби ми могли чітко встановлювати відмінності в молекулярній організації та функціонуванні організмів, ми змогли б значно краще розібратися в походженні та еволюції різних організмів, ніж на підставі морфологічних спостережень”. , Що "в основі всіх навіть суто морфологічних ознак, на підставі яких ми класифікуємо та встановлюємо види, лежать саме біохімічні відмінності"**.
* (Е. R. Lankester. Uber das Vorcommen von Haemoglobin in den Muskeln der Mollusken und die Verbreitung desselben in den lebendigen Organismen.- "Pfluger"s Archiv fur die gesammte Physiol., 1871, Bd 4, 319.)
** (В. Л. Комаров. Вибрані соч., Т. 1. М.-Л., Вид-во АН СРСР, 1945, стор 331.)
А. В. Благовіщенський і С. Л. Іванов ще в 20-х роках зробили перші в нашій країні кроки щодо з'ясування деяких питань еволюції та систематики організмів на основі порівняльного аналізу їхнього біохімічного складу (див. гл. 2). Порівняльний аналіз структури білків і нуклеїнових кислот у цей час стає все більш відчутною підмогою для систематиків (див. розділ 21). Цей метод молекулярної біології дозволяє як уточнити становище окремих видів у системі, а й змушує по-новому подивитись самі принципи класифікації організмів, котрий іноді переглянути всю систему загалом, як і сталося, наприклад, із систематикою мікроорганізмів. Безсумнівно, й у майбутньому аналіз структури геному займатиме центральне місце у хемосистематиці організмів.
Велике значення для становлення молекулярної біології мало розшифрування механізмів реплікації ДНК та транскрипції (див. розділ 24).
Біосинтез білка
Важливе зрушення у вирішенні проблеми біосинтезу білка пов'язане з успіхами у вивченні нуклеїнових кислот. У 1941 р. Т. Касперсон (Швеція) та 1942 р. Ж. Браше (Бельгія) звернули увагу на те, що в тканинах з активним білковим синтезом міститься підвищена кількість РНК. Вони дійшли висновку, що рибонуклеїнові кислоти грають визначальну роль синтезі білка. У 1953 р. Е. Гейл і Д. Фокс, начебто, отримали прямі докази безпосередньої участі РНК у біосинтезі білка: за їхніми даними, рибонуклеаза значно пригнічувала включення амінокислот у лізатах бактеріальних клітин. Аналогічні дані були отримані В. Олфрі, М. Делі та А. Мирським (1953) на гомогенатах печінки. Пізніше Еге. Гейл відмовився від висловленої їм правильної ідеї про провідну роль РНК у білковому синтезі, помилково вважаючи, що активація білкового синтезу у безклітинній системі відбувалася під впливом якоїсь іншої речовини невідомої природи. У 1954 р. П. Замітник, Д. Літлфілд, Р. Б. Хесін-Лур'є та інші виявили, що найбільш активне включення амінокислот відбувається у багатих РНК фракціях субклітинних частинок – мікросом. П. Замечник та Е. Келлер (1953 – 1954) виявили, що включення амінокислот помітно посилювалося у присутності надосадової фракції в умовах регенерації АТФ. П. Сікевіц (1952) та М. Хогланд (1956) виділили з надосадової рідини білкову фракцію (рН 5 фракція), яка була відповідальною за різке стимулювання включення амінокислот у мікросомах. Поряд із білками в надосадовій рідині було виявлено особливий клас низькомолекулярних РНК, які тепер називають транспортними РНК (тРНК). У 1958 р. Хогланд і Помічник, і навіть П. Берг, Р. Світ і Ф. Аллен та ще дослідники виявили, що з активації кожної амінокислоти необхідний свій спеціальний фермент, АТФ і специфічна тРНК. Стало ясно, що тРНК виконують виключно функцію адаптерів, тобто пристосувань, які знаходять на нуклеїновій матриці (іРНК) місце відповідної амінокислоті у білковій молекулі, що формується. Ці дослідження повністю підтвердили адапторну гіпотезу Ф. Крику (1957), що передбачала існування в клітині полінуклеотидних адапторів, необхідних для правильного розташування амінокислотних залишків білка, що синтезується, на нуклеїновій матриці. Вже багато пізніше французький учений Ф. Шапвіль (1962) в лабораторії Ф. Ліпмана (Нобелевська премія, 1953) у США дуже дотепно і однозначно показав, що місце розташування амінокислоти в білковій молекулі, що синтезується, повністю визначається тією специфічною тРНК, до якої вона приєднана. Адапторна гіпотеза Крику була розвинена у роботах Хогланда та Помічника.
До 1958 стали відомі такі основні етапи білкового синтезу: 1) активація амінокислоти специфічним ферментом з "рН 5 фракції" в присутності АТФ з утворенням аміноациладенілату; 2) приєднання активованої амінокислоти до специфічної тРНК із вивільненням аденозинмонофосфату (АМФ); 3) зв'язування аміноацил-тРНК (тРНК, навантажена амінокислотою) з мікросомами та включення амінокислот у білок з вивільненням тРНК. Хогланд (1958) зазначив, що на останньому етапі білкового синтезу необхідний гуанозінтріфосфат (ГТФ).
Транспортні РНК та синтез гена
Після виявлення тРНК почалися активні пошуки їх фракціонування та визначення нуклеотидної послідовності. Найбільших успіхів досяг американський біохімік Р. Холлі. У 1965 р. він встановив структуру аланінової тРНК із дріжджів. За допомогою рибонуклеаз (гуанілова РНК-аза і панкреатична РНК-аза) Холлі розділив молекулу нуклеїнової кислоти на кілька фрагментів, визначив у кожному окремо нуклеотидну послідовність і потім реконструював послідовність всієї молекули аланінової тРНК. Цей шлях аналізу нуклеотидної послідовності отримав назву блочного методу. Заслуга Холлі полягала головним чином тому, що він навчився розділяти молекулу РНК як на дрібні шматки, як і багато хто й до нього, а й у великі фрагменти (четвертинки і половинки). Це і дало можливість правильно зібрати окремі маленькі шматки воєдино і цим відтворити повну нуклеотидну послідовність всієї молекули тРНК (Нобелевская премія, 1968).
Цей прийом відразу ж був прийнятий на озброєння у багатьох лабораторіях світу. Протягом наступних двох років у СРСР і там була розшифрована первинна структура відразу кількох тРНК. А. А. Баєв (1967) та співробітники вперше встановили послідовність нуклеотидів у дріжджовій валіновій тРНК. До теперішнього часу вивчено вже понад десяток різних індивідуальних тРНК. Своєрідний рекорд у визначенні нуклеотидної послідовності встановлено у Кембриджі Ф. Сенгером та Г. Браунлі. Ці дослідники розробили напрочуд витончений метод поділу олігонуклеотидів і встановили послідовність так званої 5 S (рибосомної) РНК із клітин кишкової палички (1968). Ця РНК складається з 120 нуклеотидних залишків і, на відміну від тРНК, не містить додаткових мінорних основ, які помітно полегшують аналіз нуклеотидної послідовності, служачи унікальними орієнтирами окремих фрагментів молекули. В даний час завдяки використанню методу Сенгера та Браунлі успішно просувається робота з вивчення послідовності довгих рибосомних РНК та деяких вірусних РНК у лабораторії Ж. Ебеля (Франція) та інших дослідників.
А. А. Баєв та співробітники (1967) виявили, що розрізана навпіл валінова тРНК відновлює свою макромолекулярну структуру в розчині і, незважаючи на дефект у первинній структурі, має функціональну активність вихідної (нативної) молекули. Цей підхід – реконструкція розрізаної макромолекули після видалення певних фрагментів – виявився дуже перспективним. Він широко використовується зараз з'ясування функціональної ролі окремих ділянок тих чи інших тРНК.
В останні роки досягнуто великого успіху в отриманні кристалічних препаратів індивідуальних тРНК. Зараз у кількох лабораторіях у США та Англії вдалося закристалізувати вже багато тРНК. Це дало змогу досліджувати стуруктуру тРНК за допомогою рентгеноструктурного аналізу. У 1970 р. Р. Бок представив перші рентгенограми та тривимірні моделі кількох тРНК, створені ним у Вісконсінському університеті. Ці моделі допомагають визначити локалізацію окремих функціонально активних ділянок у тРНК та зрозуміти основні засади функціонування цих молекул.
Найважливіше значення для розкриття механізму синтезу білка та вирішення проблеми специфічності цього процесу мало розшифровка природи генетичного коду (див. розділ 24), яку без перебільшення можна розглядати як провідне завоювання природознавства XX ст.
Розкриття Р. Холлі первинної структури тРНК дало поштовх роботам Г. Корани* (США) щодо синтезу олігонуклеотидів і направило їх на шлях синтезу певної біологічної структури – молекули ДНК, що кодує аланінову тРНК. Зроблені Кораною майже 15 років тому перші кроки з хімічного синтезу коротких олигонуклеотидов завершилися 1970 р. вперше здійсненим синтезом гена. Корана та його співробітники спочатку з окремих нуклеотидів синтезували хімічним шляхом короткі фрагменти завдовжки 8-12 нуклеотидних залишків. Ці фрагменти із заданою нуклеотидною послідовністю утворювали спонтанно двотяжкі комплементарні шматки з перекриттям 4 - 5 нуклеотидів. Потім ці готові шматки в потрібному порядку послідовно з'єднували кінець в кінець за допомогою ферменту ДНК-лігази. Таким чином, на відміну від реплікації молекул ДНК, за А. Корнбергом** (див. розділ 24), Корані вдалося заново створити молекулу природної двотяжкової ДНК за заздалегідь наміченою програмою відповідно до послідовності тРНК, описаної Холлі. Аналогічним чином зараз ведуться роботи з синтезу інших генів (М. М. Колосов, 3. А. Шабарова, Д. Г. Кнорре, 1970 – 1975).
* (За дослідження генетичного коду Г. Корані та М. Ніренбергу була присуджена у 1968 р. Нобелівська премія.)
** (За відкриття полімерази та синтез ДНК А. Корнбергу, а за синтез РНК С. Очоа у 1959 р. було присуджено Нобелівську премію.)
Мікросоми, рибосоми, трансляція
У 1950-х років вважалося, що центром білкового синтезу у клітині є мікросоми. Термін мікросоми був вперше введений у 1949 р. А. Клодом для позначення фракції дрібних гранул. Пізніше з'ясувалося, що за білковий синтез відповідальна не вся фракція мікросом, що складається з мембран та гранул, а лише дрібні рибонуклеопротеїдні частки. Ці частки 1958 р. були названі Р. Робертсом рибосомами.
Класичні дослідження бактеріальних рибосом були проведені А. Тисьєром та Дж. Уотсоном у 1958 – 1959 рр. Бактеріальні рибосоми виявилися дещо дрібнішими за рослинні та тваринні. Дж. Літлтон (1960), М. Кларк (1964) та Е. Н. Светайло (1966) показали, що рибосоми хлоропластів вищих рослин та мітохондрій належать до бактеріального типу. А. Тисьєр та інші (1958) виявили, що рибосоми дисоціюють на дві нерівні субодиниці, що містять по одній молекулі РНК. Наприкінці 50-х років вважалося, що кожна молекула рибосомної РНК складається з кількох коротких фрагментів. Проте А. С. Спірін у 1960 р. вперше показав, що РНК у субчастинках представлені безперервною молекулою. Д. Уоллер (1960), розділивши рибосомні білки за допомогою електрофорезу в крохмальному гелі, встановив, що вони гетерогенні. Спочатку багато хто сумнівався в даних Уоллера, оскільки здавалося, що білок рибосоми повинен бути строго гомогенним, як, наприклад, білок ВТМ. В даний час в результаті досліджень Д. Уоллер, Р. Траута, П. Трауба та інших біохіміків стало відомо, що до складу власне рибосомних частинок входить більше 50 абсолютно різних за структурою білків. А. С. Спіріну в 1963 р. вдалося вперше розгорнути рибосомні субчастинки і показати, що рибосоми є компактно скрученим рибонуклеопротеїдним тяжом, який в певних умовах може розгортатися. У 1967 – 1968 рр. М. Номура повністю реконструював біологічно активну субчастицю з рибосомної РНК та білка і навіть отримав такі рибосоми, у яких білок та РНК належали різним мікроорганізмам.
До сьогодні незрозуміла роль рибосомної РНК. Передбачається, що вона є тією унікальною специфічною матрицею, на якій при формуванні рибосомної частки знаходить строго певне місце кожен із численних рибосомних білків (А. С. Спірін, 1968).
А. Річ (1962) виявив агрегати з кількох рибосом, з'єднаних між собою ниткою іРНК. Ці комплекси було названо полісомами. Виявлення полісом дозволило Річу і Уотсону (1963) висловити припущення, що синтез поліпептидного ланцюга відбувається на рибосомі, яка просувається по ланцюжку іРНК. У міру просування рибосоми по ланцюжку іРНК в частинці відбувається зчитування інформації та утворення поліпептидного ланцюга білка, а нові рибосоми по черзі приєднуються до прочитаного кінця іРНК, що вивільняється. З даних Річа та Уотсона випливало, що значення полісом у клітині полягає у масовій продукції білка шляхом послідовного прочитування матриці відразу декількома рибосомами.
В результаті досліджень М. Ніренберга, С. Очоа, Ф. Ліпмана, Г. Корани та інших у 1963 – 1970 рр. в. стало відомо, що поряд з іРНК, рибосомами, АТФ та аміноацил-тРНК у процесі трансляції бере участь велика кількість різноманітних факторів, а сам процес трансляції може бути умовно поділений на три етапи – ініціацію, власне трансляцію та термінацію.
Ініціація трансляції означає синтез першого пептидного зв'язку в комплексі рибосома – матричний полінуклеотид – аміноацил-тРНК. Таку ініціаторну активність має не всяка аміноацил-тРНК, а формілметіоніл-тРНК. Ця речовина була вперше виділена у 1964 р. Ф. Сенгером та К. Маркером. С. Бретчер і К. Маркер (1966) показали, що ініціаторна функція формілметіоніл-тРНК обумовлена її підвищеною спорідненістю до пептидильного центру рибосоми. Для початку трансляції дуже важливими є також деякі білкові фактори ініціації, які були виділені в лабораторіях С. Очоа, Ф. Гро та інших дослідницьких центрах. Після утворення першого пептидного зв'язку в рибосомі починається власне трансляція, тобто послідовне приєднання аміноацильного залишку до С-кінця поліпептиду. Багато деталей процесу трансляції вивчили К. Монро та Дж. Бішоп (Англія), І. Рихлік та Ф. Шорм (ЧССР), Ф. Ліпман, М. Бретчер, В. Гілберт (США) та інші дослідники. У 1968 р. А. С. Спірін для пояснення механізму роботи рибосоми запропонував оригінальну гіпотезу. Привідним механізмом, що забезпечує всі просторові переміщення тРНК та іРНК під час трансляції, є періодичне розмикання та змикання субчастинок рибосоми. Закінчення трансляції закодовано в матриці, що зчитується, яка містить термінуючі кодони. Як показав С. Бреннер (1965 – 1967), такими кодонами є триплети УАА, УАГ та УГА. М. Капеччі (1967) виявив також спеціальні білкові фактори термінації. А. С. Спіріним та Л. П. Гавриловою описаний так званий "неферментативний" синтез білка в рибосомах (1972 - 1975) без участі білкових факторів. Це відкриття важливе для розуміння походження та еволюції біосинтезу білка.
Регуляція активності генів та білків
Після проблеми специфічності білкового синтезу першому місці у молекулярної біології виявилася проблема регуляції синтезу білків, чи, що саме, регуляції активності генів.
Функціональна нерівнозначність клітин та пов'язані з нею репресія та активація генів давно привертали увагу генетиків, але досі реальний механізм контролю генної активності залишався невідомим.
Перші спроби пояснити регуляторну активність генів пов'язані з вивченням гістонних білків. Ще подружжя Стедман на початку 40-х років XX ст. висловлювали думку, що гістони можуть грати у цьому явищі основну роль. Надалі вони отримали перші чіткі дані про відмінності у хімічній природі гістонних білків. Нині кількість фактів, які свідчать користь цієї гіпотези, з кожним роком дедалі більше зростає.
* (Е. Stedman, E. Stedman. The basic proteins of cell nuclei.- Phylosoph. Trans. Roy. Soc. London, 1951, v. 235, 565 – 595.)
У той самий час накопичується дедалі більше даних, які говорять у тому, що регуляція генної активності - набагато складніший процес, ніж просте взаємодію ділянок генів з молекулами гістонних білків. У 1960 – 1962 рр. в лабораторії Р. Б. Хесина-Лур'є було з'ясовано, що гени фагів починають зчитуватися неодночасно: гени фага Т2 можна поділити на ранні, функціонування яких відбувалося в перші хвилини зараження бактеріальної клітини, і пізні, що починали синтезувати іРНК після завершення ранніх генів.
У 1961 р. французькі біохіміки Ф. Жакоб і Ж. Моно запропонували схему регулювання активності генів, яка зіграла виняткову роль у розумінні регуляторних механізмів клітини взагалі. Згідно зі схемою Жакоба і Моно, у ДНК крім структурних (інформаційних) генів є ще гени-регулятори та гени-оператори. Ген-регулятор кодує синтез специфічної речовини - репресора, який може приєднуватись як до індуктора, так і до гена-оператора. Ген-оператор зчеплений зі структурними генами, а ген-регулятор перебуває у певному віддаленні них. Якщо серед немає індуктора, наприклад, лактози, то синтезований геном-регулятором репресор зв'язується з геном-оператором і, блокуючи його, вимикає роботу всього оперону (блок структурних генів разом з керуючим ними оператором). Утворення ферменту цих умовах немає. Якщо ж середовищі з'являється індуктор (лактоза), то продукт гена-регулятора - репресор - пов'язується з лактозою і знімає блок з гена-оператора. В цьому випадку стає можливою роботаструктурного гена, що кодує синтез ферменту, та фермент (лактоза) утворюється в середовищі.
На думку Жакоба і Моно, ця схема регуляції застосовна до всіх адаптивних ферментів і може бути як при репресії, коли утворення ферменту пригнічується надлишком продукту реакції, і при індукції, коли внесення субстрату викликає синтез ферменту. За дослідження регулювання активності генів Жакоб і Моно були удостоєні в 1965 р. Нобелівської премії.
Спочатку ця схема здавалася надто надуманою. Однак згодом з'ясувалося, що регуляція генів за цим принципом має місце не лише у бактерій, а й у інших організмів.
Починаючи з 1960 р. помітне місце в молекулярній біології займають дослідження організації геному і структури хроматину в еукаріотичних організмів (Дж. Боннер, Р. Бріттен, В. Олфрі, П. Уокер, Ю. С. Ченцов, І. Б. Збарський та ін.) .) та з регуляції транскрипції (А. Мирський, Г. П. Георгієв, М. Бернстіл, Д. Голл, Р. Цанев, Р. І. Салганік). Довгий час залишалася невідомою та спірною природа репресора. У 1968 р. Пташне (США) показав, що репресором є білок. Він виділив його в лабораторії Дж. Уотсона і виявив, що репресор, дійсно, має спорідненість до індуктора (лактози) і одночасно "пізнає" ген-оператор лак-оперону і специфічно зв'язується з ним.
В останні 5 - 7 років отримані дані про наявність ще одного керуючого осередку генної активності - промотор. Виявилося, що по сусідству з операторною ділянкою, до якої приєднується продукт, синтезований на ген-регуляторі - білковій речовині репресора, є інша ділянка, яку також слід віднести до членів регуляторної системи генної активності. До цієї ділянки приєднується білкова молекула ферменту РНК-полімерази. У промоторному ділянці має відбутися взаємне впізнавання унікальної послідовності нуклеотидів у ДНК та специфічної конфігурації білка РНК-полімерази. Від ефективності впізнавання залежатиме здійснення процесу зчитування генетичної інформації з даною послідовністю генів оперону, що примикає до промотору.
Крім описаної Жакобом і Моно схеми, у клітині існують інші механізми регуляції генів. Ф. Жакоб та С. Бреннер (1963) встановили, що регуляція реплікації бактеріальної ДНК певним чином контролюється клітинною мембраною. Досліди Жакоба (1954) з індукції різних профагів переконливо показали, що під впливом різних мутагенних факторів у клітині лізогенних бактерій починається виборча реплікація гена профагу, а реплікація геному господаря блокується. У 1970 р. Ф. Белл повідомив у тому, що у цитоплазму з ядра можуть переходити невеликі молекули ДНК і там транскрибироваться.
Таким чином, регуляція активності генів може здійснюватися на рівні реплікації, транскрипції та трансляції.
Значних успіхів досягнуто у вивченні регуляції як синтезу ферментів, а й їх активності. На явища регуляції активності ферментів у клітині вказували ще у 50-х роках А. Новік та Л. Сциллард. Г. Умбаргер (1956) встановив, що у клітині існує дуже раціональний шлях придушення активності ферменту кінцевим продуктом ланцюга реакцій на кшталт зворотний зв'язок. Як було встановлено Ж. Моно, Ж. Шанже, Ф. Жакобом, А. Парді та іншими дослідниками (1956 – 1960), регуляція активності ферментів може здійснюватися за алостеричним принципом. Фермент або одна з його субодиниць, крім спорідненості до субстрату, має спорідненість з одним з продуктів ланцюга реакцій. Під впливом такого продукту-сигналу фермент змінює свою конформацію, що втрачає активність. В результаті весь ланцюг ферментативних реакцій вимикається на самому початку. На суттєву роль конформаційних змін білка в ферментативних реакціях, а у відомому сенсі і на наявність алостеричного ефекту, вказували Д. Вімен та Р. Вудворд (1952; лауреат Нобелівської премії, 1965).
Структура та функції білків
В результаті робіт Т. Осборна, Г. Гофмейстера, А. Гюрбера, Ф. Шульца та багатьох інших наприкінці XIXв. було отримано багато тварин та рослинні білки в кристалічному вигляді. Приблизно в цей час за допомогою різних фізичних методів були встановлені молекулярні ваги деяких білків. Так, в 1891 р. А. Сабанєєв і Н. Александров повідомили, що молекулярна вага овальбуміну становить 14 000; в 1905 р. Е. Рейд встановив, що молекулярна вага гемоглобіну дорівнює 48 000. Полімерна структура білків була розкрита в 1871 р. Г. Глазівець і Д. Габерманом. Ідея про пептидний зв'язок окремих амінокислотних залишків у білках була висловлена Т. Куртіусом (1883). Роботи з хімічної конденсації амінокислот (Е. Шаал, 1871; Г. Шифф, 1897; Л. Бальбіано та Д. Траскіатті, 1900) та синтезу гетерополіпептидів (Е. Фішер, 1902 - 1907, хімічна структура білків.
Перший кристалічний фермент (уреаза) було отримано 1926 р. Дж. Самнером (Нобелевская премія, 1946), а 1930 р. Дж. Нортроп (Нобелевская премія, 1946) отримав кристалічний пепсин. Після цих робіт стало зрозуміло, що ферменти мають білкову природу. У 1940 р. М. Куніц виділив кристалічну РНК-азу. До 1958 вже було відомо більше 100 кристалічних ферментів і понад 500 ферментів, виділених у некристалічному вигляді. Одержання високоочищених препаратів індивідуальних білків сприяло розшифровці їхньої первинної структури та макромолекулярної організації.
Велике значення у розвиток молекулярної біології взагалі і генетики людини, особливо, мало відкриття Л. Полингом (1940) ненормального гемоглобіну S, виділеного з еритроцитів людей із тяжкою спадковою хворобою - серповидно-клітинної анемією. У 1955 – 1957 рр. В. Інгрем використовував розроблений Ф. Сенгер метод "відбитків пальців" (плям, утворених окремими пептидами при хроматографії на папері) для аналізу продуктів гідролізу гемоглобіну S лугом і трипсином. У 1961 р. Інгрем повідомив, що гемоглобін S відрізняється від нормального гемоглобіну тільки за природою одного амінокислотного залишку: у нормальному гемоглобіні в сьомому положенні ланцюга знаходиться залишок глютамінової кислоти, а в гемоглобіні S - залишок валіну. Цим повністю підтвердилося (1949) припущення Полінга, що серповидно-клітинна анемія є хворобою молекулярної природи. Спадкова зміна всього одного залишку амінокислоти в кожній половинці макромолекули гемоглобіну призводить до того, що гемоглобін втрачає здатність легко розчинятися при низькій концентрації кисню і починає кристалізуватися, що призводить до порушення структури клітини. Ці дослідження з усією очевидністю показали, що структура білка є суворо певною амінокислотною послідовністю, яка закодована в геномі. Про виняткове значення первинної структури білка у формуванні унікальної біологічно активної конформації макромолекули свідчили роботи К. Анфінсена (1951). Анфінсен показав, що біологічно активна макроструктура панкреатичної рибонуклеази, що втрачається в результаті відновлення, зумовлена амінокислотною послідовністю і може знову виникати спонтанно при окисленні SH-груп залишків цистеїну з утворенням дисульфідних зшивок у строго визначених місцях пептидного ланцюга ферменту.
До теперішнього часу детально вивчений механізм дії великої кількості ферментів та визначено структуру багатьох білків.
У 1953 р. Ф. Сенгер встановив амінокислотну послідовність інсуліну. :Цей білок складається з двох поліпептидних ланцюгів, з'єднаних двома дисульфідними зшивками. Один з ланцюгів містить всього 21 амінокислотний залишок, а інший - 30 залишків. На розшифрування будівлі цього порівняно простого білка Сенгер витратив близько 10 років. У 1958 р. за це видатне дослідження йому було присуджено Нобелівську премію. Після створення В. Стейном та С. Муром (1957) автоматичного аналізатора амінокислот, ідентифікація продуктів часткового гідролізу білків значно прискорилася. У 1960 р. Стейн і Мур вже повідомили у тому. що їм вдалося визначити послідовність рибонуклеази, пептидний ланцюжок якого представлений 124 амінокислотними залишками. У тому ж році в лабораторії Г. Шрамма у Тюбінгені (ФРН) Ф. Андерер та інші визначили амінокислотну послідовність у білку ВТМ. Потім амінокислотна послідовність була визначена в міоглобіні (А. Едмунсон) та α- та β-ланцюгах гемоглобіну людини (Г. Браунітцер, Е. Шредер та ін), лізоцимі з білка курячого яйця (Ж. Жолле, Д. Кейфілд). У 1963 р. Ф. Шорм та Б. Кейл (ЧССР) встановили послідовність амінокислот у молекулі хімотрипсиногену. У тому ж році було визначено амінокислотну послідовність трипсиногену (Ф. Шорм, Д. Уолш). У 1965 р. К. Такахаші встановив первинну структуру рибонуклеази Т1. Потім послідовність амінокислот було визначено ще в кількох білків.
Як відомо, остаточним доказом правильності визначення тієї чи іншої структури є її синтез. У 1969 р. Р. Меріфілд (США) вперше здійснив хімічний синтез панкреатичної рибонуклеази. За допомогою розробленого ним методу синтезу на твердофазовому носії Меріфілд приєднував до ланцюжка одну амінокислоту за іншою відповідно до тієї послідовності, яка була описана Стейном і Муром. В результаті він отримав білок, який за своїми якостями був ідентичний панкреатичній рибонуклеазі А. За розкриття будови рибонуклеази В. Стейну, С. Муру та К. Анфінсену була у 1972 р. присуджено Нобелівську премію. Цей синтез природного білка відкриває грандіозні перспективи, вказуючи на можливість створення будь-яких білків відповідно до запланованої послідовності.
З рентгеноструктурних досліджень У. Астбері (1933) випливало, що пептидні ланцюги білкових молекул скручені або укладені якимось строго певним чином. Починаючи з цього часу, багато авторів висловлювали різні гіпотези про способи укладання білкових ланцюгів, але до 1951 всі моделі залишалися умоглядними побудовами, що не відповідали експериментальним даним. У 1951 р. Л. Полінг та Р. Корі опублікували серію блискучих робіт, у яких остаточно було сформульовано теорію вторинної структури білків - теорію α-спіралі. Поряд з цим стало також відомо, що білки мають ще третинну структуру: α-спіраль пептидного ланцюга може бути певним чином складена, утворюючи досить компактну структуру.
У 1957 р. Дж. Кендрю та його співробітники вперше запропонували тривимірну модель структури міоглобіну. Ця модель потім уточнювалася протягом декількох років, поки в 1961 не з'явилася підсумкова робота з характеристикою просторової структури цього білка. У 1959 р. М. Перутц та співробітники встановили тривимірну структуру гемоглобіну. На цю роботу дослідники витратили понад 20 років (перші рентгенограми гемоглобіну були отримані Перутцем 1937 р.). Оскільки молекула гемоглобіну складається з чотирьох субодиниць, то, розшифрувавши його організацію, Перутц цим вперше описав четвертинну структуру білка. За роботи з визначення тривимірної структури білків Кендрю та Перутцу у 1962 р. було присуджено Нобелівську премію.
Створення перуком просторової моделі структури гемоглобіну дозволило. наблизитися до розуміння механізму функціонування цього білка, який, як відомо, здійснює перенесення кисню у клітинах тварин. Ще 1937 р. Ф. Гауровиц дійшов висновку у тому, що взаємодія гемоглобіну з киснем, повітря має супроводжуватися зміною структури білка. У 60-х роках Перутц та його співробітники виявили помітне зміщення ланцюгів гемоглобіну після його окислення, що викликалося зрушенням атомів заліза внаслідок зв'язування з киснем. На цій основі сформувалися уявлення про "дихання" білкових макромолекул.
У 1960 р. Д. Філліпс та його співробітники розпочали рентгеноструктурні дослідження молекули лізоциму. До 1967 р. їм більш-менш точно вдалося встановити деталі організації цього білка та локалізацію окремих атомів у його молекулі. Крім цього, Філіпс з'ясував характер приєднання лізоциму до субстрату (тріацетилглюкозаміну). Це дозволило відтворити механізм цього ферменту. Таким чином, знання первинної структури та макромолекулярної організації дало змогу не лише встановити природу активних центрів багатьох ферментів, а й повністю розкрити механізм функціонування цих макромолекул.
Використання методів електронної мікроскопії допомогло розкрити принципи макромолекулярної організації таких складних білкових утворень, як нитки колагену, фібриногену, скорочувальних фібрил м'язів та ін. Наприкінці 50-х років було запропоновано моделі м'язового скорочувального апарату. Виняткове значення розуміння механізму м'язового скорочення мало відкриття У. А. Енгельгардтом і М. М. Любимової (1939) АТФ-азной активності міозину. Це означало, що в основі акта м'язового скорочення лежить зміна фізико-хімічних властивостей та макромолекулярної організації скоротливого білка під впливом аденозинтрифосфорної кислоти (див. розділ 11).
Для розуміння принципів складання біологічних структур важливе значення мали вірусологічні дослідження (див. Розділ 25).
Невирішені проблеми
Основні успіхи у сучасній молекулярній біології досягнуто переважно у результаті вивчення нуклеїнових кислот. Проте навіть у цій галузі ще далеко не всі проблеми вирішені. Великих зусиль вимагатиме, зокрема, розшифровка всієї нуклеотидної послідовності геному. Ця проблема у свою чергу нерозривно пов'язана з проблемою гетерогенності ДНК і вимагає розробки нових досконалих методів фракціонування та виділення індивідуальних молекул із сумарного генетичного матеріалу клітини.
До цих пір зусилля в основному були зосереджені на окремому вивченні білків та нуклеїнових кислот. У клітині ці біополімери нерозривно пов'язані один з одним і функціонують головним чином у формі нуклеопротеїдів. Тому зараз з особливою гостротою виявилася необхідність вивчення взаємодії білків та нуклеїнових кислот. На перший план висувається проблема впізнавання білками певних ділянок нуклеїнових кислот. Вже намітилися кроки до вивчення такої взаємодії цих біополімерів, без якого немислимо повне розуміння структури та функцій хромосом, рибосом та інших структур. Без цього неможливо також усвідомити регулювання активності генів і остаточно розшифрувати принципи роботи білоксинтезуючих механізмів. Після робіт Жакоба та Моно з'явилися деякі нові дані про регуляторне значення мембран у синтезі ядерного матеріалу. Це ставить завдання глибшого дослідження ролі мембран у регуляції реплікації ДНК. У цілому нині проблема регуляції активності генів і клітинної активності взагалі стала однією з найважливіших проблем сучасної молекулярної біології.
Сучасний стан біофізики
У зв'язку з проблемами молекулярної біології йшов розвиток біофізики. Інтерес до цієї галузі біології стимулювався, з одного боку, необхідністю всебічного вивчення впливу на організм різноманітних випромінювань, з іншого - потребою дослідження фізичних і фізико-хімічних основ життєвих явищ, що протікають на молекулярному рівні.
Отримання точних відомостей про молекулярні структури і процесах, що в них відбуваються, стало можливим в результаті застосування нових тонких фізико-хімічних методів. На основі досягнень електрохімії вдалося вдосконалити метод вимірювання біоелектричних потенціалів, застосувавши іонно-виборчі електроди (Г. Ейзенман, Б. П. Нікольський, Кхурі, 50 – 60-і роки). Дедалі ширше входить у практику інфрачервона спектроскопія (з використанням лазерних пристроїв), що дозволяє досліджувати конформаційні зміни білків (І. Плотніков, 1940). Цінні відомості дає також метод електронного парамагнітного резонансу (Е. К. Завойський, 1944) та біохемолюмінесцентний метод (Б. Н. Тарусов та ін., 1960), які дозволяють, зокрема, судити про транспорт електронів при окисних процесах.
До 50-х років біофізика завойовує міцне становище. Виникає потреба у підготовці кваліфікованих спеціалістів. Якщо у 1911 р. у Європі лише в університеті м. Печ, в Угорщині, була кафедра біофізики, то до 1973 р. такі кафедри існують майже у всіх великих університетах.
У 1960 р. було організовано Міжнародне товариство біофізиків. Торішнього серпня 1961 р. відбувся перший Міжнародний біофізичний конгрес у Стокгольмі. Другий конгрес був проведений у 1965 р. у Парижі, третій – у 1969 р. у Бостоні, четвертий – у 1972 р. у Москві.
У біофізиці зберігається чітке розмежування між двома різними за змістом напрямками – молекулярною біофізикою та клітинною біофізикою. Це розмежування отримує й організаційний вираз: створюються окремі кафедри цих двох напрямів біофізики. У Московському університеті перша кафедра біофізики була створена в 1953 р. на біолого-грунтовому факультеті, дещо пізніше виникла кафедра біофізики на фізичному факультеті. За таким же принципом організовувалися кафедри у багатьох інших університетах.
Молекулярна біофізика
В останні роки все більше зміцнювався зв'язок молекулярної біофізики з молекулярною біологією, і зараз іноді буває важко визначити, де проходить межа поділу між ними. У генеральному наступі на проблему спадкової інформації така кооперація біофізики з молекулярною біологією неминуча.
Головним напрямом у дослідницькій роботі є вивчення фізики нуклеїнових кислот – ДНК та РНК. Застосування зазначених вище методів і насамперед рентгеноструктурного аналізу сприяло розшифровці молекулярної структури нуклеїнових кислот. В даний час ведуться інтенсивні дослідження з вивчення поведінки цих кислот у розчинах. Особлива увага приділяється при цьому конформаційним переходам "спіраль-клубок", що вивчаються щодо змін в'язкості, оптичних та електричних показників. У зв'язку з вивченням механізмів мутагенезу розвиваються дослідження щодо вивчення дії іонізуючої радіації на поведінку нуклеїнових кислот у розчинах, а також дії радіації на нуклеїнові кислоти вірусів та фагів. Всебічного аналізу піддавався вплив ультрафіолетового випромінювання, деякі спектральні ділянки якого добре поглинаються нуклеїновими кислотами. Велику питому вагу в таких дослідженнях займає виявлення активних радикалів нуклеїнових кислот і білків методом електронного парамагнітного резонансу. Із застосуванням цього пов'язано виникнення цілого самостійного напрями.
Проблема кодування інформації ДНК та РНК та її передачі при синтезі білка давно цікавила молекулярну біофізику, і фізики неодноразово висловлювали з цього приводу ті чи інші міркування (Е. Шредінгер, Г. Гамов). Розшифровка генетичного коду викликала численні теоретичні та експериментальні дослідження по структурі спіралі ДНК, механізму ковзання та закручування її ниток, вивчення фізичних сил, що беруть участь у даних процесах.
Значну допомогу молекулярна біофізика надає молекулярної біології у вивченні структури білкових молекул з допомогою рентгеноструктурного аналізу, вперше застосованого 1930 р. Дж. Берналом. Саме в результаті використання фізичних методів у поєднанні з біохімічними (ферментативні методи) було розкрито молекулярну конформацію та послідовність розташування амінокислот у ряді білків.
Сучасні електронно-мікроскопічні дослідження, що виявили наявність у клітинах та її органоїдах складних мембранних систем, стимулювали спроби зрозуміти їхню молекулярну будову (див. глави 10 та 11). Вивчається прижиттєво хімічний склад мембран та, зокрема, властивості їх ліпідів. Було з'ясовано, що останні здатні до переокислення та неферментативних реакцій ланцюгового окиснення (Ю. А. Володимиров та Ф. Ф. Литвин, 1959; Б. Н. Тарусов та ін., 1960; І. І. Іванов, 1967), що призводить до порушення мембранних функцій. Для вивчення складу мембран стали користуватися також методами математичного моделювання (В. Ц. Пресман, 1964 – 1968; М. М. Шемякін, 1967; Ю. А. Овчинников, 1972).
Клітинна біофізика
Знаменною подією в історії біофізики стало формування в 50-х роках чітких уявлень про термодинаміку біологічних процесів, внаслідок чого остаточно відпали припущення про можливість самостійного утворення енергії в живих клітинах всупереч другому закону термодинаміки. Розуміння дії цього закону в біологічних системах пов'язане із запровадженням бельгійським ученим І. Пригожиним (1945) * у біологічну термодинаміку поняття відкритих систем, що обмінюються із зовнішнім середовищем енергією та матерією. Пригожин показав, що позитивна ентропія утворюється в живих клітинах при робочих процесах відповідно до другого закону термодинаміки. Введені ним рівняння визначили умови, у яких виникає так зване стаціонарне стан (раніше його називали також динамічним рівновагою), у якому кількість вільної енергії (негентропії), що надходить у клітини з їжею, компенсує її витрата, а позитивна ентропія виводиться. Це відкриття підкріпило загальнобіологічну ідею про нерозривний зв'язок зовнішнього та внутрішнього середовища клітин. Воно поклало початок реальному вивченню термодинаміки живих систем, у тому числі методом моделювання (А. Бертон, 1939; А. Г. Пасинський, 1967).
* (Загальну теорію відкритих систем вперше висунув Л. Берталанфі у 1932 р.)
Відповідно до основного принципу біотермодинаміки, необхідною умовою існування життя виявляється стаціонарність у розвитку її біохімічних процесів, для здійснення якої необхідна координація швидкостей численних реакцій обміну речовин. На основі нової біофізичної термодинаміки виник напрям, що виділяє зовнішні та внутрішні фактори, які забезпечують цю координацію реакцій та роблять її стійкою. За останні два десятиліття виявлено велику роль у підтримці стаціонарного стану системи інгібіторів і особливо антиоксидантів (Б. Н. Тарусов та А. І. Журавльов, 1954, 1958). Встановлено, що надійність стаціонарного розвитку пов'язана з факторами зовнішнього середовища (температурою) та фізико-хімічними властивостями середовища клітин.
Сучасні принципи біотермодинаміки дозволили надати фізико-хімічне тлумачення механізму адаптації. За нашими даними, пристосування до умов зовнішнього середовища може відбуватися тільки в тому випадку, якщо за їх зміни організм здатний встановити стаціонарність у розвитку біохімічних реакцій (Б. Н. Тарусов, 1974). Постало питання розробці нових методів, які б оцінювати стаціонарний стан прижиттєво і прогнозувати його можливі порушення. Велику користь обіцяє впровадження в біотермодинаміки та дослідження процесів біологічної адаптації кібернетичних принципів саморегулівних систем. Стало ясно, що для вирішення питання про стійкість стаціонарного стану важливий облік так званих факторів, що обурюють, до яких відносяться, зокрема, неферментативні реакції окислення ліпідів. Останнім часом дедалі більше розширюються дослідження процесів переокислення в ліпідних фазах живих клітин та наростання активних радикальних продуктів, що порушують регуляторні функції мембран. Джерелом інформації про ці процеси служить як виявлення активних перекисних радикалів, так і перекисних сполук біоліпідів (А. Таппель, 1965; І. І. Іванов, 1965; Є. Б. Бурлакова, 1967 та інші). Для виявлення радикалів використовують біохемолюмінесценцію, що виникає у ліпідах живих клітин при їх рекомбінації.
На основі фізико-хімічних уявлень про стабільність стаціонарного стану виникли біофізичні уявлення про адаптацію рослин до змін умов зовнішнього середовища як порушення інгібуючих антиокислювальних систем (Б. Н. Тарусов, Я. Є. Доскоч, Б. М. Кітлаєв, А. М. Агавердієв , 1968 – 1972). Це відкрило можливість оцінювати такі властивості, як морозостійкість та солестійкість, а також робити відповідні прогнози під час селекції сільськогосподарських рослин.
У 50-х роках було відкрито надслабке світіння – біохемолюмінесценція низки біологічних об'єктів у видимій та інфрачервоній частинах спектру (Б. Н. Тарусов, А. І. Журавльов, А. І. Поливода). Це стало можливо в результаті розробки методів реєстрації надслабких світлових потоків за допомогою фотоелектронних помножувачів (Л. А. Кубецький, 1934). Будучи результатом біохімічних реакцій, що протікають у живій клітині, біохемолюмінесценція дозволяє судити про важливі окисні процеси в ланцюгах перенесення електронів між ферментами. Відкриття та вивчення біохемолюмінесценції має велике теоретичне та практичне значення. Так, Б. Н. Тарусов та Ю. Б. Кудряшов відзначають велику роль продуктів окислення ненасичених жирних кислот у механізмі виникнення патологічних станів, що розвиваються під дією іонізуючих випромінювань, при канцерогенезі та інших порушеннях нормальних функцій клітини.
У 50-х роках у зв'язку з бурхливим розвитком ядерної фізики з біофізики виділилася радіобіологія, що досліджує біологічну дію іонізуючих випромінювань. Одержання штучних радіоактивних ізотопів, створення термоядерної зброї, атомних реакторів та розвиток інших форм практичного використання атомної енергії поставило з усією гостротою проблему захисту організмів від шкідливої дії іонізуючої радіації, розробки теоретичних основ профілактики та лікування променевої хвороби. Для цього необхідно було насамперед з'ясувати, які компоненти клітини та ланки обміну речовин є найбільш уразливими.
Об'єктом вивчення біофізики та радіобіології стало з'ясування природи первинних хімічних реакцій, що виникають у живих субстратах під впливом енергії випромінювань. Тут було важливо як зрозуміти механізми цього явища, а й зуміти впливати на процес розміни фізичної енергії на хімічну, зменшити його коефіцієнт " корисного " дії. Роботам у цьому напрямі започаткували дослідження школи Н. Н. Семенова (1933) в СРСР та Д. Хіншельвуда (1935) в Англії.
Велике місце у радіобіологічних дослідженнях зайняло вивчення ступеня радіаційної опірності різних організмів. Було встановлено, що підвищена радіорезистентність (наприклад, гризунів пустель) обумовлена високою антиокислювальною активністю ліпідів. клітинних мембран(М. Чанг та ін., 1964; Н. К. Огризов та ін., 1969). Виявилося, що у формуванні антиоксидативних властивостей цих систем велику роль відіграють токофероли, вітамін К і тіосполуки (І. І. Іванов та ін., 1972). В останні роки велику увагу привертають до себе також дослідження механізмів мутагенезу. З цією метою вивчається дія іонізуючих випромінювань на поведінку нуклеїнових кислот та білків in vitro, а також у вірусах та фагах (А. Густафсон, 1945 – 1950).
Боротьба за подальше підвищення ефективності хімічного захисту, пошук ефективніших інгібіторів та принципів інгібування залишаються у цьому напрямі основними завданнями біофізики.
Просунулося вперед дослідження збуджених станів біополімерів, що визначають їхню високу хімічну активність. Найбільш успішно йшло вивчення збуджених станів, що виникають на первинній стадії фотобіологічних процесів – фотосинтезу та зору.
Так, зроблено солідний внесок у розуміння первинної активації молекул пігментних систем рослин. Встановлено велике значення перекидання (міграції) енергії збуджених станів без втрат активованих пігментів на інші субстрати. Велику роль розвитку цих уявлень зіграли теоретичні роботи А. М. Тереніна (1947 і пізніше). А. А. Красновський (1949) відкрив і досліджував реакцію оборотного фотохімічного відновлення хлорофілу та його аналогів. Нині складається загальне переконання, що найближчим часом можна буде відтворити фотосинтез у штучних умовах (див. також розділ 5).
Біофізики продовжують працювати над розкриттям природи м'язового скорочення та механізмів нервового збудження та проведення (див. розділ 11). Актуального значення набули також дослідження механізмів переходу від збудженого стану до норми. Збуджений стан розглядають тепер як результат автокаталітичної реакції, а гальмування - як наслідок різкої мобілізації інгібіторної антиокислювальної активності в результаті молекулярних перегрупувань в таких сполуках, як токоферол (І. І. Іванов, О. Р. Кольс, 1966; О. Р. Кольс, 1970).
Найважливішою загальною проблемою біофізики залишається пізнання якісних фізико-хімічних особливостей живої матерії. Такі властивості, як здатність живих біополімерів вибірково пов'язувати калій або поляризувати електричний струм, не вдається зберегти навіть при їхньому обережному вилученні з організму. Тому клітинна біофізика продовжує інтенсивно розробляти критерії та методи для прижиттєвого дослідження живої матерії.
Попри молодість молекулярної біології, успіхи, досягнуті нею у цій галузі, воістину приголомшливі. За порівняно короткий термін встановлено природу гена та основні принципи його організації, відтворення та функціонування. Понад те, здійснено як розмноження генів in vitro, а й уперше завершено повний синтез самого гена. Повністю розшифровано генетичний код і вирішено найважливішу біологічну проблему специфічності біосинтезу білка. Виявлено та досліджено основні шляхи та механізми утворення білка в клітині. Повністю визначено первинну структуру багатьох транспортних РНК - специфічних молекул-адапторів, що здійснюють переклад мови нуклеїнових матриць на мову амінокислотної послідовності білка, що синтезується. До кінця розшифрована амінокислотна послідовність багатьох білків та встановлена просторова структура деяких з них. Це дозволило з'ясовувати принцип і деталі функціонування молекул ферментів. Здійснено хімічний синтез одного з ферментів – рибонуклеази. Встановлено основні принципи організації різних субклітинних частинок, багатьох вірусів та фагів та розгадано основні шляхи їх біогенезу у клітині. Розкрито підходи до розуміння шляхів регуляції активності генів та з'ясування регуляторних механізмів життєдіяльності. Вже простий перелік цих відкриттів свідчить у тому, що друга половина XX в. ознаменувалася величезним прогресом біології, який зобов'язаний насамперед поглибленому вивченню структури та функції біологічно найважливіших макромолекул - нуклеїнових кислот і білків.
Досягнення молекулярної біології вже сьогодні використовуються на практиці та приносять відчутні плоди у медицині, сільському господарстві та деяких галузях промисловості. Безперечно, що віддача цієї науки зростатиме з кожним днем. Однак головним підсумком все ж таки слід вважати, що під впливом успіхів молекулярної біології зміцнилася впевненість у існуванні необмежених можливостей на шляху розкриття найпотаємніших таємниць життя.
У майбутньому, мабуть, будуть відкриті нові шляхи дослідження біологічної форми руху матерії – з молекулярного рівня біологія перейде на атомарний рівень. Однак зараз не знайдеться, мабуть, жодного дослідника, який міг би реально передбачити розвиток молекулярної біології навіть на найближчі 20 років.
Молекулярна біологія / м? лɛ доJʊ лər / є гілкою біології, що стосується молекулярної основи біологічної активності між біомолекул у різних системах клітини, у тому числі взаємодій між ДНК, РНК, білків та їх біосинтезу, а також регулювання цих взаємодій. Запис у природі 1961 року, Астбері описав молекулярну біологію:
Не так техніка, як підхід, підхід з погляду про фундаментальних наук із провідною ідеєю пошуку нижче великомасштабних проявів класичної біології для відповідного молекулярного плану. Він стурбований тим, зокрема, з формамибіологічних молекул і [...] переважно тривимірним і структурно - що не означає, однак, що це лише уточнення морфології. Він має водночас досліджувати генезис та функції.
Ставлення до інших біологічних наук
Дослідники в галузі молекулярної біології використовують специфічні методи ростуть молекулярну біологію, але все більше і більше комбінують їх з методами та ідеями від генетики та біохімії. Існує не певна межа між цими дисциплінами. Це показано на наступній схемі, що зображує один можливий вид відносин між полями:
- Біохімія є вивчення хімічних речовин і життєво важливих процесів, що відбуваються в живих організмах. Біохіміки важко зосередитися на ролі, функції та структури біомолекул. Вивчення хімії за біологічних процесів та синтезу біологічно активних молекул є прикладами біохімії.
- Генетика є вивчення впливу генетичних відмінностей у організмах. Це часто може бути виведено у відсутності нормальної компоненти (наприклад один ген). Вивчення «мутанти»-організми мають один або більше функціональні компоненти по відношенню до так званого «дикого типу» або нормального фенотипу. Генетичні взаємодії (епістаз) часто плутають прості інтерпретації таких «нокаут» дослідження.
- Молекулярна біологія є вивчення молекулярних основ процесів реплікації, транскрипції, трансляції та функції клітин. Центральна догма молекулярної біології, де генетичний матеріал транскрибується в РНК і потім транслюється в білок, незважаючи на спрощене, як і раніше забезпечує хорошу початкову точку для розуміння поля. Картина була переглянута у світлі нових ролей для РНК .
Методи молекулярної біології
Молекулярне клонування
Одним із найголовніших методів молекулярної біології для вивчення функції білка є молекулярним клонуванням. У цій техніці, ДНК, що кодує білок, що представляє інтерес, клонованої за допомогою полімеразної ланцюгової реакції (ПЛР), та/або ферменти рестрикції в плазміді (вектора експресії,). Вектор має 3 відмінні риси: початок реплікації, а сайт множинного клонування (MCS), і селективний маркер, як правило, зі стійкістю до антибіотиків. Розташовані вище сайт множинного клонування є промоторними областями та транскрипції сайту ініціації, що регулюють експресію клонованого гена. Ця плазміда може бути вставлена в або бактеріальні або тваринні клітини. Введення ДНК у бактеріальні клітини може бути зроблено шляхом трансформації за допомогою поглинання голої ДНК, кон'югацій за допомогою міжклітинних контактів або трансдукції за допомогою вірусного вектора. Введення ДНК в еукаріотичні клітини, такі як клітини тварин, за допомогою фізичних чи хімічних засобів, називається трансфекцією. Декілька різних методів трансфекції доступні, такі як фосфат кальцію трансфекції, електропорації, мікроін'єкції та ліпосомальної трансфекції. Плазміда може бути інтегрована в геном, що призводить до стабільної трансфекції, або може залишатися незалежними від геному, званого перехідними процесами трансфекції.
ДНК, що кодують білки, що представляє інтерес, в даний час всередині клітини, і білки тепер можуть бути виражені. Різноманітні системи, такі як індуцибельні промотори та специфічні клітинні сигнальні фактори, які допоможуть виявити інтерес білок на високих рівнях. Великі кількості білка можуть бути витягнуті з бактеріальної або еукаріотичної клітини. Білок може бути перевірений на ферментативну активність при різних ситуаціях, білка можна кристалізувати, тому його третинна структура може бути вивчена, або, у фармацевтичній промисловості, активність нових препаратів проти білка може бути вивчена.
Полімеразної ланцюгової реакції
Макромолекули-блот та дослідження
Терміни північний , західнийі східнийБлоттінг отримує з того, що спочатку була молекулярна біологія жарт, яка грала на терміні Саузернет, після методики, описаної Edwin Southern для гібридизації BLOTTED ДНК Патриція Томас, розробник РНК - блот, який потім став відомий як північному - блоттингунасправді не використовувати цей термін.
Саузернблот
Названий на честь його винахідника, біолог Едвін Південний, то Саузерн - блот є методом дослідження на наявність специфічної послідовності ДНК у зразку ДНК. Зразки ДНК до або після ферменту рестрикції (рестриктаз) переварювань розділені за допомогою електрофорезу в гелі, а потім переносили на мембрану за допомогою блотингу за допомогою капілярної дії. Мембрану потім піддають впливу міченого ДНК - зонда, який має послідовність підстав доповненням до послідовності ДНК, що представляє інтерес. Саузерн - блоттинг менш широко використовується в науковій лабораторії через здатність інших методів, таких як ПЛР, для виявлення специфічних послідовностей ДНК із зразків ДНК. Ці блоти все ще використовуються для деяких застосувань, однак, таких як вимірювання трансгену числа копій у трансгенних мишах або в інженерії гена нокаутних ліній ембріональних стовбурових клітин.
Північний Блоттінг
Northern блот діаграма
Східно-блотингу
Клінічні дослідження та медичні методи лікування, що випливають із молекулярної біології, частково охоплені генною терапією. Застосування молекулярної біології або молекулярна клітинна біологія підходів у медицині тепер називається молекулярною медициною. Молекулярна біологія також відіграє важливу роль у розумінні освіти, дій і нормативних актів різних частин клітин, які можуть бути використані для ефективних призначати нові ліки, хвороба діагнозу та зрозуміти фізіологію клітини.
подальше читання
- Cohen, SN, Чанг, НКД, Бойєр, H. & Heling, RB Конструювання біологічно функціональних бактеріальних плазмід у пробірці .
1. Введення.
Предмет, завдання та методи молекулярної біології та генетики. Значення "класичної" генетики та генетики мікроорганізмів у становленні молекулярної біології та генної інженерії. Поняття гена в "класичній" та молекулярній генетиці, його еволюція. Внесок методології генної інженерії у розвиток молекулярної генетики. Прикладне значення генної інженерії для біотехнології.
2. Молекулярні засади спадковості.
Поняття про клітину, її макромолекулярний склад. Природа генетичного матеріалу. Історія доказу генетичної функції ДНК.
2.1. Різні види нуклеїнових кислот.Біологічні функції нуклеїнових кислот. Хімічна будова, просторова структура та фізичні властивості нуклеїнових кислот. Особливості будови генетичного матеріалу про- та еукаріотів. Комплементарні пари основ Уотсона-Кріка. генетичний код. Історія розшифрування генетичного коду. Основні властивості коду: триплетність, код без ком, виродженість. Особливості кодового словника, сім'ї кодонів, смислові та «безглузді» кодони. Кільцеві молекули ДНК та поняття про надспіралізацію ДНК. Топоізомери ДНК та їх типи. Механізми дії топоізомераз. ДНК-гіраза бактерій.
2.2. Транскрипція ДНК.РНК-полімераза прокаріотів, її субодинична та тривимірна структури. Різноманітність сигма-факторів. Промотор генів прокаріотів, його структурні елементи. Стадії транскрипційного циклу. Ініціація, освіта "відкритого комплексу", елонгація та термінація транскрипції. Атенюація транскрипції. Регулювання експресії триптофанового оперону. "Рибоперемикачі". Механізми термінації транскрипції. Негативне та позитивне регулювання транскрипції. Лактозний оперон. Регулювання транскрипції у розвитку фага лямбда. Принципи впізнавання ДНК регуляторними білками (САР-білок та репресор фага лямбда). Особливості транскрипції у еукаріотів. Процесинг РНК у еукаріотів. Кепірування, сплайсинг та поліаденілювання транскриптів. Механізми сплайсингу. Роль малих ядерних РНК та білкових факторів. Альтернативний сплайсинг, приклади.
2.3. Трансляція, її етапи, функція рибосом Локалізація рибосом у клітці. Прокаріотичний та еукаріотичний типи рибосом; 70S та 80S рибосоми. Морфологія рибосом. Підрозділ на субчастинки (субодиниці). Кодон-залежне зв'язування аміноацил-тРНК в елонгаційному циклі. Кодон-антикодонова взаємодія. Участь фактора елонгації EF1 (EF-Tu) у зв'язуванні аміноацил-тРНК із рибосомою. Чинник елонгації EF1В (EF-Ts), його функція, послідовність реакцій з його участю. Антибіотики, що впливають на етап кодон-залежного зв'язування аміноацил-тРНК із рибосомою. Аміноглікозидні антибіотики (стрептоміцин, неоміцин, канаміцин, гентаміцин та ін.), механізм їхньої дії. Тетрацикліни як інгібітори зв'язування аміноацил-тРНК із рибосомою. Ініціація трансляції. Основні етапи процесу ініціації. Ініціація трансляції у прокаріотів: фактори ініціації, ініціаторні кодони, 3¢-кінець РНК малої рибосомної субчастинки та послідовність Шайна-Дальгарно у мРНК. Ініціація трансляції у еукаріотів: фактори ініціації, ініціаторні кодони, 5¢-нетрансльована область та кеп-залежна «кінцева» ініціація. "Внутрішня" кеп-незалежна ініціація у еукаріотів. Транспептидація. Інгібітори транспептидації: хлорамфенікол, лінкоміцин, аміцетин, стрептограміни, анізоміцин. Транслокація. Участь фактора елонгації EF2 (EF-G) та ГТФ. Інгібітори транслокації: фусидова кислота, віоміцин, їх механізми дії. Термінація трансляції. Термінуючі кодони. Білкові фактори термінації прокаріотів та еукаріотів; два класи факторів термінації та механізми їх дії. Регуляція трансляції у прокаріотів.
2.4. Реплікація ДНКта її генетичний контроль. Полімерази, що беруть участь у реплікації, характеристика їх ферментативних активностей. Точність відтворення ДНК. Роль стеричних взаємодій між парами основ ДНК під час реплікації. Полімерази I, II та III E. coli. Субодиниці полімерази III. Виделка реплікації, “провідна” та “відстаюча” нитки під час реплікації. Фрагменти Оказаки. Комплекс білків у вилки реплікації. Регуляція ініціації реплікації E. соli. Термінація реплікації у мікробів. Особливості регулювання реплікації плазмід. Двонаправлена реплікація і реплікація за типом кільця, що котиться.
2.5. Рекомбінація, її типи та моделі. Загальна чи гомологічна рекомбінація. Двониткові розриви ДНК, що ініціюють рекомбінацію. Роль рекомбінації у постреплікативній репарації двониткових розривів. Структура Холлідея в моделі рекомбінації. Ензимологія загальної рекомбінації E. coli. RecBCD комплекс. RecA білок. Роль рекомбінації у забезпеченні синтезу ДНК при пошкодженнях ДНК, що переривають реплікацію. Рекомбінація у еукаріотів. Ферменти рекомбінації у еукаріотів. Сайт-специфічна рекомбінація. Відмінності молекулярних механізмів загальної та сайт-специфічної рекомбінації. Класифікація рекомбіназу. Типи хромосомних перебудов, що здійснюються при сайт-специфічній рекомбінації. Регуляторна роль сайт-специфічної рекомбінації у бактерій. Конструювання хромосом багатоклітинних еукаріотів за допомогою системи сайт-специфічної рекомбінації фага.
2.6. Репарація ДНК.Класифікація типів репарації. Пряма репарація тімінових димерів та метильованого гуаніну. Вирізання основ. Глікозилази. Механізм репарації неспарених нуклеотидів (mismatch репарація). Вибір нитки ДНК, що репарується. SOS-репарація. Властивості ДНК полімераз, що беруть участь у SOS-репарації у прокаріотів та еукаріотів. Уявлення про "адаптивні мутації" у бактерій. Репарація двониткових розривів: гомологічна постреплікативна рекомбінація та поєднання негомологічних кінців молекули ДНК. Взаємозв'язок процесів реплікації, рекомбінації та репарації.
3. Мутаційний процес.
Роль біохімічних мутантів у формуванні теорії один ген – один фермент. Класифікація мутацій. Точкові мутації та хромосомні перебудови, механізм їх утворення. Спонтанний та індукований мутагенез. Класифікація мутагенів. Молекулярний механізм мутагенезу. Взаємозв'язок мутагенезу та репарації. Ідентифікація та селекція мутантів. Супресія: внутрішньогенна, міжгенна та фенотипічна.
4. Позахромосомні генетичні елементи.
Плазміди, їх будова та класифікація. Статевий фактор F, його будова та життєвий цикл. Роль фактора F у мобілізації хромосомного перенесення. Утворення донорів типу Hfr і F". Механізм кон'югації. Бактеріофаги, їх структура та життєвий цикл. Вірулентні та помірні бактеріофаги. Лізогенія та трансдукція. Загальна та специфічна трансдукція. Мігруючі генетичні елементи: транспозони та IS-послідовність. Транспозони в геномах прокаріотів та еукаріотів IS-послідовності бактерій, їх структура IS-послідовності як компонент F-фактору бактерій, що визначає здатність передачі генетичного матеріалу при кон'югації Транспозони бактерій та еукаріотичних організмів переносі транспозонів та його ролі у структурних перебудовах (эктопическая рекомбінація) й у еволюції геному.
5. Дослідження структури та функції гена.
Елементи генетичного аналізу. Цис-транс комплементаційний тест. Генетичне картування з використанням кон'югації, трансдукції та трансформації. Побудова генетичних карток. Тонке генетичне картування. Фізичний аналіз структури гена. Гетеродуплексний аналіз. Рестрикційний аналіз. Методи секвенування. Полімеразна ланцюгова реакція. Виявлення функції гена.
6. Регулювання експресії генів. Концепції оперону та регулону. Контроль лише на рівні ініціації транскрипції. Промотор, оператор та регуляторні білки. Позитивний та негативний контроль експресії генів. Контроль лише на рівні термінації транскрипції. Катаболіт-контрольовані оперони: моделі лактозного, галактозного, арабінозного та мальтозного оперонів. Атенюатор-контрольовані оперони: модель триптофанового оперону. Мультивалентне регулювання експресії генів. Глобальні системи регулювання. Регуляторна відповідь на стрес. Посттранскрипційний контроль. Сигальна трансдукція. Регуляція з участю РНК: малі РНК, сенсорні РНК.
7. Основи генної інженерії. Ферменти рестрикції та модифікації. Виділення та клонування генів. Вектор для молекулярного клонування. Принципи конструювання рекомбінантних ДНК та їх введення у реципієнтні клітини. Прикладні аспекти генної інженерії.
а). Основна література:
1. Вотсон Дж., Туз Дж., Рекомбінантні ДНК: Короткий курс. - М.: Світ, 1986.
2. Гени. - М.: Світ. 1987.
3. Молекулярна біологія: структура та біосинтез нуклеїнових кислот. / За ред. . - М. Вища шк. 1990.
4. , – Молекулярна біотехнологія. М. 2002.
5. Спірін рибосоми та біосинтез білка. - М.: вища школа, 1986.
б). Додаткова література:
1. Хесін геному. - М: Наука. 1984.
2. Рибчин генетичної інженерії. - СПб.: СПбГТУ. 1999.
3. Патрушев генів. - М: Наука, 2000.
4. Сучасна мікробіологія. Прокаріоти (у 2-х тт.). - М.: Світ, 2005.
5. М. Сінгер, П. Берг. Гени та геноми. - М.: Світ, 1998.
6. Щелкунова інженерія. - Новосибірськ: З-во Сиб. Унів., 2004.
7. Степанов біологія. Структура та функції білків. - М: В. Ш., 1996.
інтерв'ю
Пирогов Сергій – учасник підготовки до олімпіади з біології, організованої "Слон та Жираф" у 2012 р.
Переможець міжнародної універсіади з біології
Переможець олімпіади «Ломоносів»
Призер регіонального етапу Всеросійської олімпіади з біології у 2012 р.
Вчиться у МДУ ім. М.В. Ломоносова на біологічному факультеті: кафедра молекулярної біології, на 6 курсі. Працює у лабораторії біохімічної генетики тварин Інституту молекулярної генетики.
- Сергію, якщо у читачів з'являться питання, вони зможуть їх тобі поставити?
Так, звичайно, поставити запитання можна хоч одразу. У цьому полі:
Натисніть, щоб поставити запитання.

- Давай почнемо зі школи, у тебе начебто була не суперкрута школа?
Я навчався у дуже слабкій московській школі школі, такій середньостатистичній ЗОШ. Щоправда, у нас була чудова вчителька з МХК, завдяки якій у нас з'явилася багато в чому номінальна "мистецтвознавча" спрямованість школи.
– А що з біологією?
Біологію у нас вела дуже літня, глуховата і різка жінка, яку всі побоювалися. Але любові до її предмета не додавало. Я ж з дитинства був захоплений біологією, років із п'яти. Читав все сам, переважно захоплюючись анатомією та зоологією. Отже, шкільні предмети існували паралельно моїм власним інтересам. Усі змінили олімпіади.
- Розкажи про це докладніше.
У 7 класі я вперше взяв участь у муніципальному етапі (звичайно, відразу майже з усіх предметів, оскільки був єдиним учнем, якого вчителі мали підстави відправити). І став переможцем із біології. Тоді школа поставилася до цього як до кумедного, але не надто цікавого факту.

- Чи це допомогло тобі в школі?
Пам'ятаю, що незважаючи на блискуче навчання, нерідко отримував від викладача з біології четвірки з причіпками на кшталт "на малюнку розрізу цибулини коріння повинні бути розфарбовані коричневим, а не сірим". Все це було досить обтяжливим. У 8 класі я знову пішов на олімпіади, але з біології мене чомусь не відправили. Натомість став переможцем та призером з інших предметів.
- А що було у 9 класі?
У 9-му класі не пішов на окружний етап. Там я несподівано набрав слабенький, прикордонний бал, який виявився все ж таки прохідним на регіональний етап. Це мало потужну мотивуючу силу - усвідомлення того, як багато виявляється я не знаю і як багато людей, які все це знають (скільки таких людей у масштабі країни я навіть боявся уявити).

– Розкажи, як ти готувався.
Інтенсивні самостійні заняття, набіги на книгарні і тисячі торішніх завдань мали цілющий ефект. Я набрав один із найбільших балів за теорію (що теж було для мене зовсім раптовим), пройшов на практичний етап... і провалив його. Тоді я ще взагалі не знав про існування практичного етапу.
- Чи вплинула на тебе олімпіада?
Моє життя радикально змінилося. Я дізнався про багато інших олімпіад, особливо полюбив ШПВ. Згодом на багатьох показував хороші результати, деякі вигравав, завдяки "Ломоносівській" отримав право на вступ без іспитів. Паралельно я вигравав олімпіади з історії мистецтва, до якого нерівно дихаю й досі. Щоправда, з практичними турами так і не дружив. В 11 класі я таки дійшов до заключного етапу, але Фортуна не була прихильною і цього разу я не встиг заповнити матрицю відповідей теоретичного етапу. Зате це дозволило вже не турбуватися за практичний.
- Ти з багатьма олімпіадниками познайомився?
Так, досі вважаю, що мені дуже пощастило з колом моїх однолітків, які значно розширили мій кругозір. Інший бік олімпіад, крім мотивації більш гармонійно вивчати предмет, було знайомство з олімпіадниками. Вже в той час я помітив, що горизонтальне спілкування часом корисніше від вертикального - з викладачами на зборах.

- Як ти вступав до ВНЗ? Вибирав факультет?
Після 11 класу я вступив до біофаку МДУ. Саме більшість моїх тодішніх товаришів зробили вибір на користь ФББ, але тут першочергову роль відіграло те, що я не став призером всеросу. Значить мені треба було б складати внутрішній іспит з математики, а в ній, особливо шкільній – вищу я полюбив значно більше – я був не сильний. І в школі була дуже слабка підготовка (нас навіть не готували до майже всієї частини). У плані інтересів вже тоді я здогадувався, що, зрештою, можна дійти будь-якого результату, незалежно від місця надходження. Згодом з'ясувалося, що багато є і випускників ФББ, які переходили в переважно мокру біологію, і навпаки - багато хороших біоінформатиків починали любителями. Хоча в той момент мені і здавалося, що на біофаку контингент буде не в приклад слабшим за ФББшний. У цьому я, безперечно, помилявся.
А ви знали?
цікаво
А ви знали?
цікаво
У таборі Слон та Жираф є зміни з біохімії та молекулярної біології, де школярі разом із досвідченими викладачі з МДУ ставлять експерименти, а також готуються до олімпіад.© Інтерв'ю брав Решетов Денис. Фотографії люб'язно надав Пирогов Сергій.




